
盐碱胁迫下燕麦叶片代谢组差异分析
以燕麦(Avena sativa)叶片为材料,结合广靶代谢组和靶向代谢组检测方法,分析了盐胁迫和碱胁迫条件下的主要差异代谢物及其代谢途径。结果发现:盐胁迫和碱胁迫条件下,燕麦叶片的主要差异代谢物均以脂类代谢物为主,有机酸是盐胁迫与碱胁迫之间的主要差异代谢物;富集分析表明,碱胁迫下的三羧酸循环(tricarboxylic acid cycle, TCA)代谢途径与盐胁迫条件下存在差别。通过检测TCA循环相关的有机酸,发现碱胁迫下燕麦叶片的柠檬酸、乌头酸、琥珀酸、延胡索酸、苹果酸、草酰乙酸含量高于盐胁迫,其中乌头酸含量明显高于其他有机酸。总之,TCA循环是盐胁迫和碱胁迫之间的主要差异代谢通路,燕麦叶片通过积累有机酸响应碱胁迫。本研究通过代谢组学方法首次证实了燕麦叶片响应盐、碱胁迫的主要差异代谢途径是TCA途径,推测乌头酸在燕麦叶片响应碱胁迫的有机酸调节过程中发挥主要作用,为解析燕麦响应盐、碱胁迫的差异机制奠定基础。
English
-
磷是生物圈重要的生命元素,也是诸多陆地生态系统的主要限制元素[1]。磷元素的生物地球化学循环过程决定了其生物有效活性以及持续供应的能力,影响着高寒生态系统的功能和结构[2]。亚高山森林-高山草地生态系统界面的复杂性、极强的敏感性和脆弱性特征,使其在高海拔生态系统中面临近年来剧烈的气候变化和逐渐加强的人类活动干扰所带来的威胁[3]。这种生态系统界面不是固定以物理或地形特征的变化为标志,而是从一个生物群过渡到另一个生物群,它的边界有着较高的物种丰富度,且在时空上是可变的[4]。高寒生态系统由于生境条件限制了树木生长致使植被低矮,出现的明显边缘地带称作高山树线[5],此生物地理模式往往代表了地貌类型位置和空间结构的变化[6]。磷的有效性是影响高山树线交错带形成及其生物地球化学循环的重要因素之一[7],植物和微生物在高山土壤出现营养限制时竞争激烈,较低的磷养分有效性抑制了树线物种的生长[8]。土壤磷多种形态的相互转化受生物地球化学作用的影响,各磷素组分的有效性及其在土壤内的迁移转化能力不同[9]。此外,磷通过影响植被生长调控碳、氮循环,研究磷的动态对于高山生物地球化学的解析十分重要[10]。Drollinger等[11]研究喜马拉雅山脉南坡的高山树线交错带发现,土壤全磷与海拔高度相关性最强,其次是植被凋落物和分解层。青藏高原东部海拔2 900~4 000 m草地的土壤养分研究发现,不同草地类型的土壤全磷含量不同,同一草地类型在不同剖面上土壤全磷的分布也有所差异[12]。高山地区普遍存在低温和水分限制,因此探讨磷的有效性的动态特征和空间格局对于认识高寒生态系统的净初级生产力及其他功能尤为重要。
青藏高原作为地球的“第三极” [13],物种的个体反应、物种间的相互作用和气候变化期间植被的时滞效应调控着亚高山森林-高山草地生态系统界面对外部干扰的响应[14],对全球生物地化循环具有重要的警示作用。土壤无机磷以多种形态存在,是上述高寒生态系统有效磷的重要来源。亚高山森林-高山草地生态系统界面的植被类型通过影响土壤性质和凋落物产量导致无机磷组分在海拔梯度上存在差异[15]。植物进入生长季,吸收土壤磷的同时植物根系也分泌一定的酸和酶,土壤中磷形态得到转化和释放[16],影响无机磷活性。季节性积雪覆盖的土壤在早春植物生长初期冻结-融化频繁发生,也会影响土壤无机磷形态的构成[17]。因此,了解土壤无机磷组分季节动态和空间分布特征以及影响因子对于理解磷循环、提高土壤磷的有效性及其利用效率具有重要意义。青藏高原岷江上游的林线交错带非生长季植被类型下的土壤无机磷的主要存在形态是钙结合态磷[18]。川西亚高山地区的研究发现,经不同恢复途径形成的森林类型显著影响土壤生物有效磷含量,而且土壤酸碱度、钙以及可溶性有机碳含量显著改变了次生粗枝云杉(Picea asperata)阔叶混交林的有效磷含量[19]。然而,针对高山地区尤其是从亚高山森林-高山草地过渡带中的土壤无机磷组分时空变化及影响因子的综合研究仍不多见。
青藏高原东缘的岷江源区的亚高山森林-高山草地生态系统界面作为高原林区到草甸区的典型过渡地带,也是研究高山地区土壤磷的理想区域[20]。为研究高寒生态系统土壤无机磷赋存特征和影响因子,本研究选择岷江源区卡卡山和斗鸡台两处亚高山森林-高山草地生态系统界面的4 条植被带(高寒草甸、树种线、树线、密闭森林)展开研究,分别在不同生长时期(生长初期、生长盛期、生长末期)和两层土壤剖面下(0-15和15-30 cm)进行取样,探讨了高寒生态系统界面土壤无机磷组分含量时空分布的异质性,进一步明确了生长期、植被带类型以及土壤深度对土壤无机磷的影响,以及其他土壤养分与土壤无机磷组分含量的关联性,便于更好地理解高寒生态系统的土壤磷循环及其有效性,并为深入研究提供野外案例支持和相关数据补充。
1. 材料与方法
1.1 研究区概况
研究区位于四川省松潘县章腊乡境内的卡卡山(32°59′03″ N,103°40′12″ E)和斗鸡台(33°02′31″ N,103°43′59″ E),地处青藏高原的岷山山脉东段,与岷江发源地弓杠岭相隔仅数公里,坐落于四川九黄旅游线路大环线,植物群落内的主要建群种基本一致。研究区域海拔3 600~4 000 m,地形复杂,受典型山地季风气候影响,冬长且无夏,雪被期长,四时不分,年均气温5~9 ℃,年均降水量700~800 mm [21]。岷江冷杉(Abies faxoniana)是斗鸡台海拔3 600 m密闭森林植被带中群落乔木层的主要优势物种,伴生有青海云杉(Picea crassifolia)、祁连圆柏(Sabina przewalskii);卡卡山由岷江冷杉和祁连圆柏形成较为单一的乔木层,有少量紫果云杉(Picea purpurea)和方枝柏(Sabina saltuaria),郁闭度较小[22]。树线生态交错带3 700~3 750 m主要由乔木岷江冷杉和灌木雪山杜鹃(Rhododendron aganniphum)、高山柳(Salix taiwanalpina)形成[18]。树种线样带3 800~3 850 m的高山灌丛以高山杜鹃(Rhododendron lapponicum)为主。卡卡山研究区最高海拔3 997 m以下高寒草甸植物种类多样,主要有高山绣线菊(Spiraea alpina)、窄叶鲜卑花(Sibiraea angustata)、金露梅(Potentilla fruticosa)、红花绿绒蒿(Meconopsis punicea)和蒿草(Artemisia) [23]。土壤类型以山地暗棕壤为主,卡卡山草甸取样处土壤为黑色草毡土,斗鸡台密闭森林土壤为棕色针叶林土,pH均呈酸性[24]。两处研究样地卡卡山树线交错带和斗鸡台树线交错带分别位于东南坡向与西北坡向,均处于国道G544旁(图1)。
![]() 图 1 研究区域两处样地不同植被带示意图Figure 1. Schematic diagram of the different vegetation zones at the two study sites
图 1 研究区域两处样地不同植被带示意图Figure 1. Schematic diagram of the different vegetation zones at the two study sites1.2 样品采集
依据前人经验和已有研究结果,在卡卡山和斗鸡台选取草甸(3 900~4 000 m)、树种线(3 800~3 850 m)、树线(3 700~3 750 m)和密闭森林(3 600 m) 4条植被带(图1),分别采集植物生长初期(6月)、生长盛期(8月)和生长末期(10月)植被下两层土壤剖面(0-15和15-30 cm)的土样。每条植被带随机选取3个3 m × 3 m的样方,样方两两间隔大于3 m。每个采样点的选择尽量位于多个树木或者灌丛的中间位置,并对多个样点的混合样品进行指标的测定。每一样方内随机选择3个取样点,环刀法取得土样后同一样方内3个同土层土样混合成1个样本放入预先编号的自封袋,带回实验室进行下一步操作,最后共取得土样144个(2处山体 × 3个生长时期 × 4条植被带 × 2层土壤深度 × 3个重复)。
1.3 测定项目及方法
土壤P分级测定采用Tiessen磷素分级法[25]。Tiessen分级法是在Hedley分级法[26]的基础上去除微生物磷和团聚体磷的测定。本研究省略了树脂交换态磷的提取,用0.5 mol·L−1 NaHCO3溶液(有效磷)、0.1 mol·L−1 NaOH溶液、1 mol·L−1 HCl逐级振荡浸提,分别测定其活性无机磷(NaHCO3-Pi)、中等活性无机磷(NaOH-Pi)、HCl-Pi含量。全磷(total phosphorus, TP)含量采用HNO3-H2O2微波消解-钼锑抗比色法[27]测定。残留无机磷(Residual-P)含量为全磷含量减去NaHCO3-P、NaOH-P和HCl-P含量。
土壤含水量(soil variable water content, VWC)采用烘干法测定;土壤酸碱度由METTLER TOLEDO pH 计测定(水 ꞉ 土 = 2.5 ꞉ 1);土壤有机碳(soil organic carbon, SOC)含量用重铬酸钾外加热法测定[28];全氮(total nitrogen, TN)含量用元素分析仪(Vario EL Ⅲ, Elementar, Langenselbold, Germany)测定。
1.4 数据处理
土壤数据进行分析前,利用卡方(K-S)检验分析数据正态性,非正态时进行数据转换,对方差同质性进行Levene检验。采用单因素方差分析法(One-way ANOVA)研究由不同生长期和植被类型引起的土壤无机磷组分含量的差异,方差齐性时采用最小显著差异法(least significance difference, LSD,α = 0.05)进行多重比较,方差不齐则使用Dunnett T3进行多重比较。通过一般线性模型(general linear model, GLM)进行多因素方差分析(multi-way ANOVA),研究采样地、生长时期、植被带类型和土壤深度及其交互作用对土壤无机磷组分含量的影响。为明确土壤无机磷形态的时空分布格局,使用冗余分析(redundancy analysis, RDA)分析土壤无机磷组分在不同生长时期与环境因子之间的关系。为进一步测试土壤无机磷组分是否受土壤生境的影响,利用线性函数进行相关分析。统计分析和作图在SPSS 22.0、Origin 2021和Canoco 5.0中完成。
2. 结果与分析
2.1 季节变化与空间格局下土壤无机磷组分特征
卡卡山和斗鸡台的土壤无机磷含量分别为17.6~659.5和41.7~570.8 mg·kg−1。4个因素中离均差平方和(sum of squares of deviation from mean, SS)反映主要变异源因子,绝对值愈大则表明这一因素对土壤无机磷组分的影响作用愈大,生长时期和植被带类型及其交互作用对不同土壤无机磷形态的影响显著(P < 0.01),且生长时期是主要的影响因素,植被带类型对土壤无机磷组分的影响次之。研究地点对NaOH-Pi、HCl-Pi和Residual-P的含量有显著差异(P < 0.05)。NaHCO3-Pi、NaOH-Pi和HCl-Pi的含量在各土壤深度间无显著差异(P > 0.05),且土壤深度与采样地、生长时期和植被带类型并无交互作用,以下将不再考虑土层深度的影响(表1)。
表 1 卡卡山和斗鸡台土壤无机磷组分含量的多因素方差分析Table 1. Multi-way ANOVA on the content of soil inorganic phosphorus fractions at Kaka Mountain and Doujitai Mountain影响因素
Impact factor活性无机磷
NaHCO3-Pi中等活性无机磷
NaOH-PiHCl-Pi 残留无机磷
Residual-PSS F P SS F P SS F P SS F P 采样地 Sampling sites (S) 0.00 0.33 0.57 0.02 5.35 0.02* 0.01 43.22 0.00** 0.28 14.98 0.00** 生长时期 Growth period (G) 0.01 30.44 0.00** 2.52 375.80 0.00** 0.06 88.51 0.00** 3.90 105.69 0.00** 植被带类型 Vegetation type (V) 0.00 4.14 0.01** 0.11 10.71 0.00** 0.04 45.98 0.00** 1.47 26.48 0.00** 土壤深度 Soil depth (D) 0.00 2.62 0.11 0.00 0.70 0.41 0.00 1.23 0.27 0.09 5.12 0.03* 采样地 × 生长时期 S × G 0.01 14.95 0.00** 0.06 8.82 0.00** 0.01 9.13 0.00** 0.26 6.96 0.00** 采样地 × 植被带类型 S × V 0.00 2.82 0.04* 0.02 1.91 0.13 0.03 26.35 0.00** 0.32 5.77 0.00** 采样地 × 土壤深度 S × D 0.00 0.02 0.89 0.01 2.42 0.12 0.00 1.63 0.20 0.16 8.76 0.00** 生长时期 × 植被带类型 G × V 0.00 2.64 0.02* 0.12 5.82 0.00** 0.01 3.29 0.01** 0.87 7.85 0.00** 生长时期 × 土壤深度 G × D 0.00 1.63 0.20 0.01 1.77 0.18 0.00 1.50 0.23 0.13 3.51 0.03* 植被带类型 × 土壤深度 V × D 0.00 1.12 0.34 0.01 1.43 0.24 0.00 0.22 0.88 0.09 1.55 0.21 采样地 × 生长时期 ×
植被带类型 S × G × V0.00 2.21 0.05* 0.04 2.17 0.05 0.01 4.72 0.00** 0.83 7.54 0.00** 采样地 × 生长时期 ×
土壤深度 S × G × D0.00 0.88 0.42 0.01 2.02 0.14 0.00 0.80 0.45 0.01 0.38 0.68 采样地 × 植被带类型 ×
土壤深度 S × V × D0.00 1.56 0.20 0.00 0.46 0.71 0.00 0.60 0.61 0.06 1.15 0.33 生长时期 × 植被带类型 ×
土壤深度 G × V × D0.00 1.93 0.08 0.03 1.39 0.23 0.00 0.20 0.98 0.13 1.15 0.34 采样地 × 生长时期 × 植被带类型 ×
土壤深度 S × G × V × D0.00 1.54 0.17 0.01 0.31 0.93 0.00 0.40 0.88 0.13 1.21 0.31 SS值为离均差平方和。* P < 0.05;** P < 0.01。
SS is the sum of squares of deviation from the mean. * P < 0.05; ** P < 0.01.卡卡山和斗鸡台不同生长期的土壤无机磷含量差别很大,尤其是NaOH-Pi,在生长期间均呈现逐渐增加的趋势并在生长末期含量达到最高(P < 0.05)。卡卡山草甸区、树种线和密闭森林处生长末期(13.5~99.8 mg·kg−1)的NaHCO3-Pi含量显著高于生长初期和生长盛期(10.7~58.5 mg·kg−1) (P < 0.05)。斗鸡台密闭森林处的生长初期(26.2 mg·kg−1) NaHCO3-Pi含量显著低于生长盛期(P < 0.05)。卡卡山各植被带(13.3~62.6 mg·kg−1)和斗鸡台草甸、树线和树种线处(25.2~114.5 mg·kg−1)的HCl-Pi含量均在生长初期最低。Residual-P在无机磷含量中占比相对较高,卡卡山各植被带(398.6~1 508.8 mg·kg−1)和斗鸡台草甸、树种线和密闭森林(81.8~782.1 mg·kg−1)的Residual-P含量在生长初期最高(图2)。
![]() 图 2 生长时期和植被带类型对卡卡山(左)和斗鸡台(右)土壤无机磷组分的影响不同小写字母表示不同生长期间差异显著(P < 0.05)。AM:草甸;TSL:树种线;TL:树线;CF:密闭森林。图3同。Figure 2. Effects of growth period and vegetation belt types on soil inorganic phosphorus fractions in Kaka Mountain (left) and Doujitai Mountain (right)Different lowercase letters mean significant differences among different growth periods at the 0.05 level. AM: meadow; TSL: tree species line; TL: tree-line; CF: closed Forest. This is applicable for Figure 3 as well.
图 2 生长时期和植被带类型对卡卡山(左)和斗鸡台(右)土壤无机磷组分的影响不同小写字母表示不同生长期间差异显著(P < 0.05)。AM:草甸;TSL:树种线;TL:树线;CF:密闭森林。图3同。Figure 2. Effects of growth period and vegetation belt types on soil inorganic phosphorus fractions in Kaka Mountain (left) and Doujitai Mountain (right)Different lowercase letters mean significant differences among different growth periods at the 0.05 level. AM: meadow; TSL: tree species line; TL: tree-line; CF: closed Forest. This is applicable for Figure 3 as well.2.2 生长季土壤无机磷形态在不同植被带的分布格局
在卡卡山,土壤无机磷组分生长初期、生长盛期和生长末期在第1轴、第2轴解释量分别为99.54%和0.27%、99.84%和0.78%、66.19%和30.40%。生长初期草甸(meadow, AM)土壤无机磷的主要形态为NaHCO3-Pi和Residual-P,树种线(tree species line, TSL)和密闭森林(closed forest, CF)土壤无机磷的主要形态为HCl-Pi和NaOH-Pi。在植物生长盛期,AM土壤无机磷的主要形态为Residual-P,CF土壤无机磷的主要形态为NaHCO3-Pi和HCl-Pi。在生长末期,AM土壤无机磷的主要形态为NaOH-Pi和Residual-P,TSL土壤无机磷的主要形态为HCl-Pi,CF土壤无机磷的主要形态为NaHCO3-Pi (图3)。
![]() 图 3 卡卡山和斗鸡台土壤无机磷形态季节动态和空间格局的冗余分析第1轴和第2轴分别代表第1主成分和第2主成分。磷形态箭头和植被带类型之间的角度越小则磷形态和该植被带之间有良好相关性。Figure 3. Redundancy analysis of seasonal dynamics and spatial distribution pattern of soil inorganic phosphorus forms at Kaka Mountain and Doujitai MountainThe first and second axes represent the first and second principal components, respectively. The smaller the angle between the arrows of a phosphorus form and a vegetation belt type, the better is the correlation between them.
图 3 卡卡山和斗鸡台土壤无机磷形态季节动态和空间格局的冗余分析第1轴和第2轴分别代表第1主成分和第2主成分。磷形态箭头和植被带类型之间的角度越小则磷形态和该植被带之间有良好相关性。Figure 3. Redundancy analysis of seasonal dynamics and spatial distribution pattern of soil inorganic phosphorus forms at Kaka Mountain and Doujitai MountainThe first and second axes represent the first and second principal components, respectively. The smaller the angle between the arrows of a phosphorus form and a vegetation belt type, the better is the correlation between them.在斗鸡台,土壤无机磷组分生长初期、生长盛期和生长末期在第1轴、第2轴解释量分别为97.07%和2.00%、94.31%和4.99%、65.97%和30.50%。植物生长初期AM土壤无机磷的主要形态为NaOH-Pi和Residual-P,TSL土壤无机磷的主要形态为HCl-Pi,CF土壤无机磷的主要形态为NaHCO3-Pi。生长盛期AM土壤无机磷的主要形态为NaOH-Pi,TSL土壤无机磷的主要形态为HCl-Pi。在生长末期,AM土壤无机磷的主要形态为NaOH-Pi和Residual-P,TSL土壤无机磷的主要形态为HCl-Pi,树线(tree-line, TL)土壤无机磷的主要形态为NaHCO3-Pi (图3)。
总体上看,在生长期内的同一样带的土壤无机磷形态较为类似,AM土壤无机磷的主要形态是中等活性无机磷(NaOH-Pi)和残留无机磷(Residual-P),TSL的主要无机磷形态是HCl-Pi,CF的主要无机磷形态是活性无机磷(NaHCO3-Pi) (图3)。
2.3 土壤无机磷形态与土壤关键理化性质的关系
土壤关键理化性质SOC、TN、TP、VWC、pH与土壤无机磷组分含量相关性显著(P < 0.05)。土壤NaHCO3-Pi含量受土壤碳含量的影响显著(P < 0.01),表现出明显的负相关关系,与土壤酸碱度呈显著正相关关系(P < 0.05)。NaOH-Pi含量与土壤全氮含量呈显著负相关关系(P < 0.01),与土壤含水量呈显著负相关关系(P < 0.05)。土壤HCl-Pi含量与土壤酸碱度呈显著正相关关系(P < 0.01)。Residual-P含量与土壤含水量呈显著正相关关系(P < 0.01) (图4)。
![]() 图 4 卡卡山和斗鸡台土壤有机碳、全氮、全磷、土壤含水量、pH与土壤无机磷组分的散点相关图有机碳、全氮、全磷、土壤含水量的样本量为144,pH的样本量为48。黑线是土壤理化性质和无机磷组分显著相关的拟合直线;* P < 0.05;** P < 0.01;图5同。Figure 4. Scatter correlation diagram between soil organic carbon, total nitrogen, total phosphorus, variable water content, pH, and soil inorganic phosphorus fractions at Kaka Mountain and Doujitai Mountainsoil organic carbon, total nitrogen, total phosphorus, variable water content sample size N = 144, pH sample size N = 48. The black line is a fitting line which has a significant correlation between soil physical and chemical properties and inorganic phosphorus fractions; * P < 0.05; ** P < 0.01. This is applicable for Figure 5 as well.
图 4 卡卡山和斗鸡台土壤有机碳、全氮、全磷、土壤含水量、pH与土壤无机磷组分的散点相关图有机碳、全氮、全磷、土壤含水量的样本量为144,pH的样本量为48。黑线是土壤理化性质和无机磷组分显著相关的拟合直线;* P < 0.05;** P < 0.01;图5同。Figure 4. Scatter correlation diagram between soil organic carbon, total nitrogen, total phosphorus, variable water content, pH, and soil inorganic phosphorus fractions at Kaka Mountain and Doujitai Mountainsoil organic carbon, total nitrogen, total phosphorus, variable water content sample size N = 144, pH sample size N = 48. The black line is a fitting line which has a significant correlation between soil physical and chemical properties and inorganic phosphorus fractions; * P < 0.05; ** P < 0.01. This is applicable for Figure 5 as well.碳氮比对土壤无机磷组分含量存在显著影响(P < 0.05)。随着碳氮比值的增加,NaHCO3-Pi和HCl-Pi含量逐渐降低(P < 0.05),土壤NaOH-Pi含量逐渐增加(P < 0.01)。NaHCO3-Pi和Residual-P含量与碳磷比呈显著负相关关系(P < 0.05)。氮磷比与Residual-P含量呈显著负相关关系(P < 0.01) (图5)。
![]() 图 5 卡卡山和斗鸡台土壤C/N、C/P、N/P与土壤无机磷组分的散点相关图Figure 5. Scatter correlation diagram of soil C/N, C/P, N/P, and inorganic phosphorus fractions at Kaka Mountain and Doujitai Mountain
图 5 卡卡山和斗鸡台土壤C/N、C/P、N/P与土壤无机磷组分的散点相关图Figure 5. Scatter correlation diagram of soil C/N, C/P, N/P, and inorganic phosphorus fractions at Kaka Mountain and Doujitai Mountain3. 讨论
3.1 生长期和植被类型对土壤无机磷组分及形态的影响
生长时期、植被带类型、土壤理化性质及其交互作用影响亚高山森林-高山草地生态系统界面的土壤无机磷形态、含量和有效性,这与前人研究结果一致[16, 29]。卡卡山和斗鸡台的土壤无机磷组分含量在不同生长时期和垂直植被带上有显著差异,生长期间的差异更甚;在土层0-15和15-30 cm几乎没有显著差异。本研究中生长时期对于NaOH-Pi和Residual-P的SS值分别为2.52和3.90,相较其他无机磷的SS值明显较大,说明生长期对这两种无机磷组分含量的影响尤为明显。Residual-P主要是由抗风化能力强的硅酸盐包裹的难以利用的无机磷,在青藏高原东缘的贡嘎山燕子沟的研究表明,当土壤pH低至4.14时,Residual-P的溶解也十分困难,因此在研究区域海拔2 334~3 600 m处Residual-P始终是土壤全磷的最大组分[30];关于未经干扰的天然次生林土壤磷素形态的垂直分布特征研究指出,在含磷较低的土壤中,较长的时间尺度内残留磷通过土壤的生物化学反应转化为有效磷[30-31]。在本研究中,残留无机磷明显较其他无机磷形态含量多,是无机磷的第一大组分,在不同植被带生长期的储量水平存在差异,且在生长初期时的含量显著最高。理县米亚罗自然保护区高山树线交错带枯枝落叶分解的研究表明,近年来气候变化背景下的林线上移近一步使植物功能群的凋落物在分解过程中加速了土壤中铁、铝元素的释放和累积[32]。本研究酸性土壤环境令有机磷矿化释放的无机磷优先被土壤中金属氧化物所固定,磷酸盐易与土壤凋落物带来的大量游离的铁铝矿物吸附结合[33]。NaOH-Pi是铁铝氧化物或氢氧化物吸附磷,铁铝氧化物或氢氧化物随土壤水分含量减少和氧化作用的加强而增加,从而提高对磷的吸附[34]。本研究中卡卡山和斗鸡台在植物生长季由于受到降雨-温度耦合以及各植被带水土保持和蒸散发作用的影响[24],造成土壤水分含量从生长初期至生长末期持续降低[卡卡山(34.3% ± 4.1%)~(45.9% ± 1.4%);斗鸡台(35.9% ± 1.7%)~(52.5% ± 2.4%)]。因此生长期内土壤的氧化作用加强,铁铝矿物对磷的吸附能力提高,NaOH-Pi含量呈现持续增加的趋势。伴随着植物刚进入生长季开始吸收土壤养分,土壤活性无机磷含量较非生长季一般有所下降[35],各植被带的NaHCO3-Pi含量在生长初期最低。植物生长期产生的凋落物包括叶片残留物通常是土壤活性磷的重要来源[36],植物进入生长盛期后,土壤微生物活性增强,凋落物增加,凋落物矿化后可以补充土壤NaHCO3-Pi含量[18]。卡卡山植物生长末期草甸区、树种线和密闭森林的NaHCO3-Pi含量最高,斗鸡台密闭森林的NaHCO3-Pi在生长盛期、生长末期显著高于生长初期,草甸样带生长末期的NaHCO3-Pi含量最高。一方面生长盛期和生长末期土壤根系分泌物的输入和凋落物的矿化增加了SOC含量[卡卡山3条样带生长盛期(3.8 ± 1.4)~(11.6 ± 2.5) g·kg−1,生长末期(10.6 ± 2.3)~(35.4 ± 8.7) g·kg−1;斗鸡台草甸生长盛期(8.6 ± 1.1) g·kg−1,生长末期(48.4 ± 4.6) g·kg−1;密闭森林样带各生长期(80.6 ± 5.9) g·kg−1,(34.3 ± 2.5) g·kg−1,(121.2 ± 9.1) g·kg−1],为土壤微生物带来能源[37-38]。另一方面土壤凋落物C/N通过对微生物数量和活性的影响调节土壤有机物质分解,促进微生物群落规模的扩大[39]。这增强了土壤酸性磷酸酶的活性,从而增加生物有效性最高的土壤无机磷含量。
生长期,以蒿草等为优势种的草甸区土壤无机磷形态主要是NaOH-Pi和Residual-P,以高山杜鹃和以岷江冷杉为优势种的树种线和密闭森林植被带的土壤无机磷形态主要是HCl-Pi和NaHCO3-Pi。在卡卡山和斗鸡台的垂直梯度上,随着海拔的上升,植被从森林过渡到灌丛和草甸,植被带土壤养分、微生物活性和凋落物质量均在发生变化[17]。植物的根系吸收无机磷、土壤微生物活化矿物磷和土壤性质变化引起无机磷的再分配等过程造成不同植被下土壤无机磷形态的差异[18]。草甸区特别是卡卡山草甸的磷形态以Residual-P为主,表明该植被带下土壤的磷活性较差。植物种类通过控制土壤pH的变化对于HCl-Pi的垂直格局分布有着十分直接的影响[15],本研究中HCl-Pi含量与pH呈现显著的正相关关系(R = 0.70,P < 0.01),树种线样带更为明显(R = 0.92,P < 0.01)。植物群落组成关系到土壤中金属元素的释放,树种线植被带土壤的HCl-Pi含量也与该植被带土壤钙含量有关[40]。Shen等[32]发现高寒生态系统以高山杜鹃等为优势种的落叶灌木凋落物中Ca元素等的释放速率显著高于常绿乔木和灌木林下凋落物,而Al和Fe在不同植物功能群下凋落物中积累,已有卡卡山的研究表明NaOH-Pi含量与Fe含量呈显著正相关关系[18]。斗鸡台生长初期和卡卡山生长盛期、生长末期密闭森林样带的主要无机磷形态是NaHCO3-Pi,这是因为针叶林的酸性土壤环境[18]和TN、C/N、N/P、VWC等因子共同控制[41]活化HCl-Pi向NaHCO3-Pi转化。
3.2 土壤关键理化性质与土壤无机磷组分的关系
在亚高山森林-高山草地生态系统界面不同植被类型下土壤理化性质影响土壤磷素形态及其循环过程。卡卡山、斗鸡台的高山灌丛和乔木林处的土壤无机磷组分与土壤关键理化性质(有机碳、全氮、全磷、含水量、pH)的相关性显著,这与前人研究结果[30, 42]相似,土壤有机质、酸性磷酸酶、土壤温度、可溶性有机碳和微生物量氮等影响土壤无机磷的有效性。小兴安岭的阔叶红松林土壤TP与各磷组分(HCl-Pi、NaHCO3-Pi和NaOH-Pi等)之间相关性显著,且所占比例较大的组分与TP相关性较高[43]。本研究中Residual-P含量明显高于HCl-Pi,与TP相关性也更为显著。土壤含水量影响土壤中的氧化还原反应,直接关系到土壤无机磷的吸附与释放[44]。Zhang等[45]发现干旱显著增加了土壤次生铁铝矿物相关的土壤无机磷含量,磷酸钙中无机磷含量显著降低,干旱条件下土壤pH降低促进磷酸钙对磷的增溶作用,而这部分释放出的磷与土壤中丰富的可交换铝通过化学反应转化为次生矿物无机磷组分。土壤含水量影响铁、铝元素与无机磷的结合,本研究中土壤含水量与NaOH-Pi含量呈显著负相关关系,这与前人研究结果一致[34]。pH较低时会增加土壤中磷酸盐的溶解度,pH为6~7时,磷的有效性最高[46]。本研究结果中,在pH为4.17~6.99时,土壤NaHCO3-Pi和HCl-Pi含量与pH呈正比,随pH的增大NaHCO3-Pi和HCl-Pi含量增加。
土壤C/N、C/P和N/P是用来衡量土壤限制性营养的重要指标[39],直接或间接地影响土壤磷素组分含量。土壤化学计量比的时空变化格局受生物环境的影响复杂[47],本研究中碳氮磷化学计量比随植被生长期、土层深度、植被样带和纬度等的变化趋势是复杂多变的,在各生长期和植被带等C/N、C/P和N/P与无机磷组分的相关关系存在差异,特别是活性磷增多的时期和样带。但是从整体上分析,土壤无机磷组分与碳氮磷化学计量比有明显的相关关系。土壤C/N通过改变微生物的群落组成和活性,对土壤磷酸酶活性产生影响[39]。本研究随着土壤C/N增大,土壤NaHCO3-Pi含量减小,NaOH-Pi含量增加,HCl-Pi含量减小。C/P反映出有机质矿化时释放P潜力的大小,是土壤P有效性的表征参数[48]。本研究中土壤无机磷活性与C/P显著相关。陆地生态系统的临界N/P作为养分限制指标来指示氮限制或磷限制,可以反映磷活性,N/P较高时土壤磷活性较低,N/P较低时则磷活性较高[49],本研究中Residual-P含量与N/P呈负相关关系。
4. 结论
植物生长期和植被带类型通过改变亚高山森林-高山草地生态系统界面土壤理化性质影响无机磷有效性。土壤的碳、氮、磷含量及其化学计量比,土壤酸碱度和含水量与土壤无机磷组分含量显著相关。土壤无机磷含量受不同海拔的植被带类型的影响,一定程度上可能也与乔灌草凋落物有关,使得土壤无机磷组分含量在生长期不同阶段的特征差异明显。
参考文献
[1] 杜良宗, 谭昆, 周少梁. 土壤盐碱地改良的研究分析与发展概况. 土壤科学, 2021, 9(1): 41-45. DU L Z, TAN K, ZHOU S L. Research analysis and development of soil saline-alkali land improvement. Hans Journal of Soil Science, 2021, 9(1): 41-45.
[2] 杨劲松. 中国盐渍土研究的发展历程与展望. 土壤学报, 2008(5): 837-845. YANG J S. Development course and prospect of saline soil research in China. Journal of Soil Science, 2008(5): 837-845.
[3] 徐璐, 王志春, 赵长巍, 王明明, 马红媛. 东北地区盐碱土及耕作改良研究进展. 中国农学通报, 2011, 27(27): 23-31. XU L, WANG Z C, ZHAO C W, WANG M M, MA H Y. Research progress of saline-alkali soil and tillage improvement in Northeast China. China Agriculture Bulletin, 2011, 27(27): 23-31.
[4] 刘丽娟, 李小玉. 干旱区土壤盐分积累过程研究进展. 生态学杂志, 2019, 38(3): 891-898. LIU L J, LI X Y. Progress on soil salt accumulation process in arid areas. Journal of Ecology, 2019, 38(3): 891-898.
[5] 燕辉, 彭晓邦, 薛建杰. NaCl 胁迫对花棒叶片光合特性及游离氨基酸代谢的影响. 应用生态学报, 2012, 23(7): 1790-1796. YAN H, PENG X B, XUE J J. Effects of NaCl stress on leaf photosynthesis characteristics and free amino acid metabolism of Heyedysarum scoparium. Journal of Applied Ecology, 2012, 23(7): 1790-1796.
[6] 郭瑞, 周际, 杨帆, 李峰. 碱胁迫对小麦(Triticum aestivum Linn.)叶片代谢过程的影响. 中国农业科学, 2017, 50(2): 250-259. GUO R, ZHOU J, YANG F, LI F. Effect of alkali stress on the leaf metabolic process of wheat (Triticum aestivum Linn.). Agricultural Science of China, 2017, 50(2): 250-259.
[7] 石德成, 盛艳敏, 赵可夫. 复杂盐碱条件对向日葵胁迫作用主导因素的实验确定. 作物学报, 2002, 28(4): 461-467. SHI D C, SHENG Y M, ZHAO K F. Experexperimental determination of the dominant factors of sunflower stress by complex salinity conditions. Journal of Crop Sciences, 2002, 28(4): 461-467.
[8] 杨春武, 李长有, 尹红娟, 鞠淼, 石德成. 小冰麦(Triticum aestivum-Agropyron intermedium)对盐胁迫和碱胁迫的生理响应. 作物学报, 2007, 33(8): 37-49. YANG C W, LI C Y, YIN H J, JU M, SHI D C. Physiological response of small ice wheat (Triticum aestivum-Agropyron intermedium) to salt stress and alkali stress. Journal of Crop Sciences, 2007, 33(8): 37-49.
[9] 麻莹, 曲冰冰, 郭立泉, 石德成, 王德利. 盐碱混合胁迫下抗碱盐生植物碱地肤的生长及其茎叶中溶质积累特点. 草业学报, 2007, 16(4): 25-33. MA Y, QU B B, GUO L Q, SHI D C, WANG D L. Growth of alkali-resistant plants under mixed stress and the characteristics of solute accumulation in stems and leaves. Acta Prataculturae Sinica, 2007, 16(4): 25-33.
[10] DU B, KRUSE J, WINKLER J B. Metabolic responses of date palm (Phoenix dactylifera L.) leaves to drought differ in summer and winter climate. Tree Physiology, 2021, 41(9): 1685-1700. doi: 10.1093/treephys/tpab027
[11] CAI S, WU L, WANG G. DA-6 improves sunflower seed vigor under Al3+ stress by regulating Al3+ balance and ethylene metabolic. Ecotoxicology and Environmental Safety, 2022, 229: 113048. doi: 10.1016/j.ecoenv.2021.113048
[12] ROMHELD V. AWAS F. Significance of root exudates in acquisition of heavy metals from a contaminated calcareous soil by graminaceous species. Journal of Plant Nutrition, 2000, 23: 1875-1866.
[13] ENRIQUE P, LUIS J, CORCUERA. JOSE M. Spatial and temporal variation in citrate and malate exudation and tissue concentration as affected by P stress in roots of white lupin. Plant and Soil, 2002, 241: 209-221. doi: 10.1023/A:1016148222687
[14] 麻莹, 王晓苹, 姜海波, 石德成. 盐碱胁迫下碱地肤体内的有机酸积累及其草酸代谢特点. 草业学报, 2017, 26(7): 158-165. MA Y, WANG X P, JIANG H B, SHI D C. Organic acid accumulation and oxalate metabolism under saline-alkali stress. Acta Prataculturae Sinica, 2017, 26(7): 158-165.
[15] 郭立泉, 陈建欣, 崔景军, 韩丹, 石德成. 盐、碱胁迫下星星草有机酸代谢调节的比较研究. 东北师大学报, 2009, 41(4): 123-128. GUO L Q, CHEN J X, CUI J J, HAN D, SHI D C. Comparative study on the metabolic regulation of star-herb organic acids under salt and alkali stress. Journal of Northeast Normal University, 2009, 41(4): 123-128.
[16] 邹春雷. 盐碱胁迫对甜菜根茎的生理代谢影响. 石河子: 第17届作物生理学术研讨会, 2022. ZOU C L. Effect of saline stress on the physiological metabolism of sugar beet. Shihezi: Abstract of the 17th Crop Physiology Society, 2022.
[17] 戴凌燕, 唐呈瑞, 殷奎德, 阮燕晔, 张立军. 苏打盐碱胁迫对甜高粱植株有机酸含量的影响. 生态学杂志, 2015, 34(3): 681-687. DAI L Y, TANG C R, YIN K D, RUAN Y Y, ZHANG L J. Effect of soda saline stress on organic acid content in sweet sorghum plants. Journal of Ecology, 2015, 34(3): 681-687.
[18] 李俊伟, 刘景辉, 王俊英, 郭来春, 王春龙, 任长忠. 盐与碱胁迫对燕麦离子平衡和有机酸含量的影响. 西北植物学报, 2022, 42(10): 1700-1710. LI J W, LIU J H, WANG J Y, GUO L C, WANG C L, REN C Z. Effect of salt and alkali stress on ionic balance and organic acid content in oat. Journal of Northwest Botany, 2022, 42(10): 1700-1710.
[19] PANG Y Y, YAN H H, GUO L C. Reference genome assemblies reveal the origin and evolution of allohexaploid oat. Nature Genetics, 2022, 54: 1248-1258. doi: 10.1038/s41588-022-01127-7
[20] 景芳, 任生兰, 边芳, 刘彦明, 张成君. 16个燕麦品种在甘肃陇中地区的适应性评价与筛选. 中国草地学报, 2023, 45(1): 68-76. JING F, REN S L, BIAN F, LIU Y M, ZHANG C J. Adaptation evaluation and screening of 16 oat varieties in Longzhong Region, Gansu Province. The Journal of the Chinese Grassland, 2023, 45(1): 68-76.
[21] 王波, 宋凤斌. 燕麦对盐碱胁迫的反应和适应性. 生态环境, 2006, 6(3): 625-629. WANG B, SOMG F B. Oat response and adaptation to saline stress. Ecological nvironment, 2006, 6(3): 625-629.
[22] 杨莉, 赵桂琴, 周向睿, 柴继宽, 杜文盼. 燕麦K+, Na+积累与AsSOS1基因表达对盐胁迫的响应. 草地学报, 2023, 31(2): 337-348. YANG L, ZHAO G Q, ZHOU X R, CHAI J K, DU W P. Oat K+, Na+ accumulation and AsSOS1 gene expression in response to salt stress. Journal of Grassland, 2023, 31(2): 337-348.
[23] 李英丽, 方正, 毛明艳. 不同燕麦品种耐碱性筛选和鉴定. 河北农业大学学报, 2014, 11(6): 17-21. LI Y L, FANG Z, MAO M Y. Basic resistance screening and identification of different oat varieties. Journal of Agricultural University of Hebei, 2014, 11(6): 17-21.
[24] ZHAO Y, LU Z, HE L. Effects of saline-alkaline stress on seed germination and seedling growth of Sorghum bicolor (L.) Moench. Appl Biochem Biotechnol, 2014, 173 (7): 1680-91.
[25] 萨如拉, 刘景辉, 刘伟, 白健慧, 王占海. 燕麦对碱胁迫的阳离子响应机制. 作物学报, 2014, 40(2): 362-368. Sarula, LIU J H, LIU W, BAI J H, WANG Z H. Cationic response mechanism of oat to alkali stress. Journal of Crop Sciences, 2014, 40(2): 362-368.
[26] 刘建新, 刘瑞瑞, 刘秀丽. 盐碱胁迫下外源硫化氢对裸燕麦叶片氨基酸代谢过程的影响. 草业学报, 2023, 32(2): 119-130. LIU J X, LIU R R, LIU X L. Effects of exogenous hydrogen sulfide on amino acid metabolism in naked oat leaves under saline-alkali stress. Acta Prataculturae Sinica, 2023, 32(2): 119-130.
[27] 刘杰, 张美丽, 张义, 石德成. 人工模拟盐、碱环境对向日葵种子萌发及幼苗生长的影响. 作物学报, 2008, 6(10): 1818-1825. doi: 10.3321/j.issn:0496-3490.2008.10.019 LIU J, ZHANG M L, ZHANG Y, SHI D C. The effect of salt and alkali environment on seed germination and seedling growth of sunflower. Crop Journal, 2008, 6(10): 1818-1825. doi: 10.3321/j.issn:0496-3490.2008.10.019
[28] BAI J H, LIU J H, ZHANG N, YANG J H, SA R L, WU L. Effect of alkali stress on soluble sugar, antioxidant enzymes and yield of oat. Journal of Integrative Agriculture, 2013, 12 (8): 1441-1449.
[29] 邹琦. 植物生理学实验指导. 北京: 中国农业出版社, 2003. ZOU Q. Experimental Auidance in Plant Physiology. Beijing: China Agricultural Press, 2003.
[30] 郭家鑫, 鲁晓宇, 陶一凡, 郭慧娟, 闵伟. 棉花在盐碱胁迫下代谢产物及通路的分析. 作物学报, 2022, 48(8): 2100-2114. GUO J X, LU X Y, TAO Y F, GUO H J, MIN W. Analysis of metabolites and pathways in cotton under saline-alkali stress. Journal of Crop Sciences, 2022, 48(8): 2100-2114.
[31] ABBASI, JAMIL, HAQ, ALI, AHMAD, MALIK. Salt stress manifestation on plants, mechanism of salt tolerance and potassium role in alleviating it: a review. Zemdirbyste, 2016, 103(2): 229-238. doi: 10.13080/z-a.2016.103.030
[32] YANG C, CHONG J, LI C, KIM C, SHI D, WANG D. Osmotic adjustment and ion balance traits of an alkali resistant halophyte Kochia sieversiana during adaptation to salt and alkali conditions. Plant Soil, 2007, 294: 263-276. doi: 10.1007/s11104-007-9251-3
[33] FOUGERE F, RUDULIER D L, STREETER J G. Effects of salt stress on amino acid, organic acid, and carbohydrate composition of roots, Bacteroids, and cytosol of alfalfa (Medicago sativa L.). Plant Physiology, 1991, 964: 1228-1236.
[34] BROSCHE M, VINOCUR B, ALATALO E R, LAMMINMAKI A, TEICHMANN T, OTTOW E A, DJILIANWV D, AFIF D, BOGEAT T M B, ALTMAN A, POLLE A, DREVER E, RUDD S, PAULIN L, AUVINEN P, KANGASJARVI J. Gene expression and metabolite profiling of Populus euphratica growing in the Negev desert. Genome Biology, 2005, 6: 681-689.
[35] 王宇超, 王得祥, 彭少兵, 何帆, 余鸽, 王小兰. 盐胁迫对木本滨藜植物细胞膜透性及生理特性的影响. 干旱地区农业研究, 2007, 25(4): 225-229. WANG Y C, WANG D X, PENG S B, HE F, YU G, WANG X L. Effect of salt stress on cell membrane permeability and physiological characteristics of S. benina plants. Agricultural Research in Arid Areas, 2007, 25(4): 225-229.
[36] 栗露露, 殷文超, 牛梅, 孟文静, 张晓星, 童红宁. 油菜素甾醇调控水稻盐胁迫应答的作用研究. 植物学报, 2019(2): 211-219. LI L L, YIN W C, NIU M, MENG W J, ZHANG X X, TONG H X. Role of brassinosterol in regulating salt stress response in rice. Journal of Botany, 2019(2): 211-219.
[37] SCHINDLER M, HAWTHOME F C. A bond-valence approach to the structure, chemistry and paragenesis of hydroxy-hydrated oxysalt minerals. ii. crystal structure and chemical composition of borate minerals. Can Mineral, 2001, 39 (5): 1243-1256.
[38] 王佳敏. 植物中ABA和JA对盐胁迫的响应. 生物过程, 2023, 13(1): 33-38. WANG J M. ABA and JA responses to salt stress in plants. Biological Processes, 2023, 13(1): 33-38.
[39] CORTI E, FALSINI S, SCHIFF S, TANI C, GONNELLI C, ALESSIO P. Saline stress impairs lipid storage mobilization during germination in Eruca sativa. Plants, 2023, 12(2): 366-366. doi: 10.3390/plants12020366
[40] LI Y X, LIU B Y, PENG Y X, LIU C L, ZHANG X Z, ZHANG Z J, LIANG W, MA F W, LI C Y. Exogenous GABA alleviates alkaline stress in Malus hupehensis by regulating the accumulation of organic acids. Scientia Horticulturae, 2020, 261: 108982.
[41] JIA X M, WANG H, SVETLA S, ZHU Y F, HU Y, CHENG L, ZHAO T, WANG Y X. Comparative physiological responses and adaptive strategies of apple Malus halliana to salt, alkali and saline-alkali stress. Scientia Horticulturae, 2019, 245: 154-162. doi: 10.1016/j.scienta.2018.10.017
[42] XING G, MA W, GAO S, JIN Z, YUE Q, YAO Y. Transcriptomic and phosphoproteomic profiling and metabolite analyses reveal the mechanism of NaHCO3 induced organic acid secretion in grapevine roots. BMC Plant Biology, 2019, 19(1): 383. doi: 10.1186/s12870-019-1990-9
[43] 石德成, 盛艳敏, 赵可夫. 复杂盐碱条件对向日葵胁迫作用主导因素的实验确定. 作物学报, 2002, 5(4): 461-467. SHI D C, SHENG Y M, ZHAO K F. Experexperimental determination of the dominant factors of sunflower stress by complex salinity conditions. Journal of Crop Sciences, 2002, 5(4): 461-467.
[44] GHOULAM C, FOURSY A, FARES K. Effects of salt stress on growth, inorganic ions and proline acumulation in relation to osmotic adjustment in five sugar et cultivars. Environmental and Experimental Botany, 2002, 47: 39-50. doi: 10.1016/S0098-8472(01)00109-5
[45] SHORT D C, COLMER T D. Salt tolerance in the halophyte Halosarcia pergranulata subsp. pergranulata. Annals of Botany, 1999, 83: 207-213. doi: 10.1006/anbo.1998.0812
[46] 杨春武, 李长有, 张美丽, 石德成. 盐、碱胁迫下小冰麦体内的pH及离子平衡. 应用生态学报, 2008, 8(5): 1000-1005. YANG C W, LI C Y, ZHANG M L, SHI D C. The pH and ion balance in small ice wheat under salt and alkali stress. Journal of Applied Ecology, 2008, 8(5): 1000-1005.
[47] 焦德志, 赵泽龙. 盐碱胁迫对植物形态和生理生化影响及植物响应的研究进展. 江苏农业科学, 2019, 47(20): 1-4. JIAO D Z, ZHAO Z L. Progress in studying the effects of salinity stress on plant morphology, physiology and biochemistry and plant response. Jiangsu Agricultural Science, 2019, 47(20): 1-4.
[48] YANG C, ZHENG S, HUANG H. Comparison of osmotic adjustment and ion balance strategies in nineteen alkali-tolerant halophyte species during adaptation to salt-alkalinized. Australian Journal of Crop Science, 2012, 6 (1): 141-148.
[49] 高玉刚. 基于转录组和代谢组联合分析燕麦响应盐碱胁迫的机制研究. 大庆: 黑龙江八一农垦大学博士学位论文, 2022. GAO Y G. Study on the responses of ota to saline-alkali stress based on transcriptome and metabolomejoint analysisis. PhD Thesis. Daqing: Heilongjiang Bayi Agricultural ReclamationUniversity, 2022.
[50] 吕家强, 李长有, 杨春武, 胡锐. 天然盐碱土壤对虎尾草茎叶有机酸积累影响及胁迫因子分析. 草业学报, 2015, 24(4): 95-103. LYU J Q, LI C Y, YANG C W, HU R. Effect of natural saline-alkali soil on organic acid accumulation and analysis of stress factors. Acta Prataculturae Sinica, 2015, 24(4): 95-103.
[51] 殷立娟, 祝玲. 羊草苗对盐碱胁迫的反应和适应性. 东北师大学报: 自然科学版, 1989, 5(4): 87-95. YIN L J, ZHU L. Response and adaptability of grass seedlings to saline stress. Journal of Northeast Normal University: Natural Science, 1989, 5(4): 87-95.
[52] 柯梅侯, 钰荣, 张一弓. 碱胁迫下冰草根系pH值与有机酸含量变化. 新疆农业科学, 2021, 58(10): 1929-1937. KE M H, YU R, ZHANG Y G. Changes of pH and organic acid content in ice roots under alkali stress. Xinjiang Agricultural Science, 2021, 58(10): 1929-1937.
[53] 郭瑞, 周际, 杨帆, 李峰, 李昊如, 夏旭, 刘琪. 拔节孕穗期小麦干旱胁迫下生长代谢变化规律. 植物生态学报, 2016, 40(12): 1319-1327. GUO R, ZHOU J, YANG F, LI F, LI H R, XIA X, LIU Q. Changes of growth metabolism under drought stress in wheat during jointing and booting stage. Journal of Plant Ecology, 2016, 40(12): 1319-1327.
-
![]()
图 1 盐、碱胁迫对燕麦地上部鲜重及叶片生理指标的影响
SA,碱胁迫处理;SS,盐胁迫处理;CK,对照。不同小写字母表示不同处理间差异显著 (P < 0.05)。图8同。
Figure 1. Effect of salt and alkali stress on oat aboveground fresh biomass and leaf physiological indexes
SA, alkali stress; SS, salt stress; CK, control. Different lowercase letters indicate significant difference between different treatments at the 0.05 level. This is applicable for Figure 8 as well.
![]()
图 2 盐碱胁迫下主成分分析
Figure 2. Principal component analysis (PCA) under salt and alkali stress
![]()
图 3 盐碱胁迫下的差异代谢物火山图
a:盐胁迫;b:碱胁迫。FC表示两组样品间表达量均值的比值。散点越大VIP值越大。红色代表显著上调的代谢物,蓝色代表显著下调的代谢物,灰色代表非显著差异的代谢物。
Figure 3. Volcano plot for SS vs CK group and SA vs CK group
a: salt stress; b: alkali stress. FC represents the ratio of the mean expression between the two samples. The larger the scatter points, the greater the VIP-value. Metabolites that are significantly up-regulated are shown in red, metabolites that are significantly down-regulated are shown in blue, and metabolites that are not significantly different are shown in gray.
![]()
图 4 盐胁迫下燕麦叶片的差异代谢物热聚类分析
Figure 4. Thermal cluster analysis of differentially accumulated metabolites in oat leaves under salt stress
![]()
图 5 碱胁迫下燕麦叶片的差异代谢物热聚类分析
Figure 5. Thermal cluster analysis of differentially accumulated metabolites in oat leaves under alkali stress
![]()
图 6 盐、碱胁迫条件下的差异代谢物分析
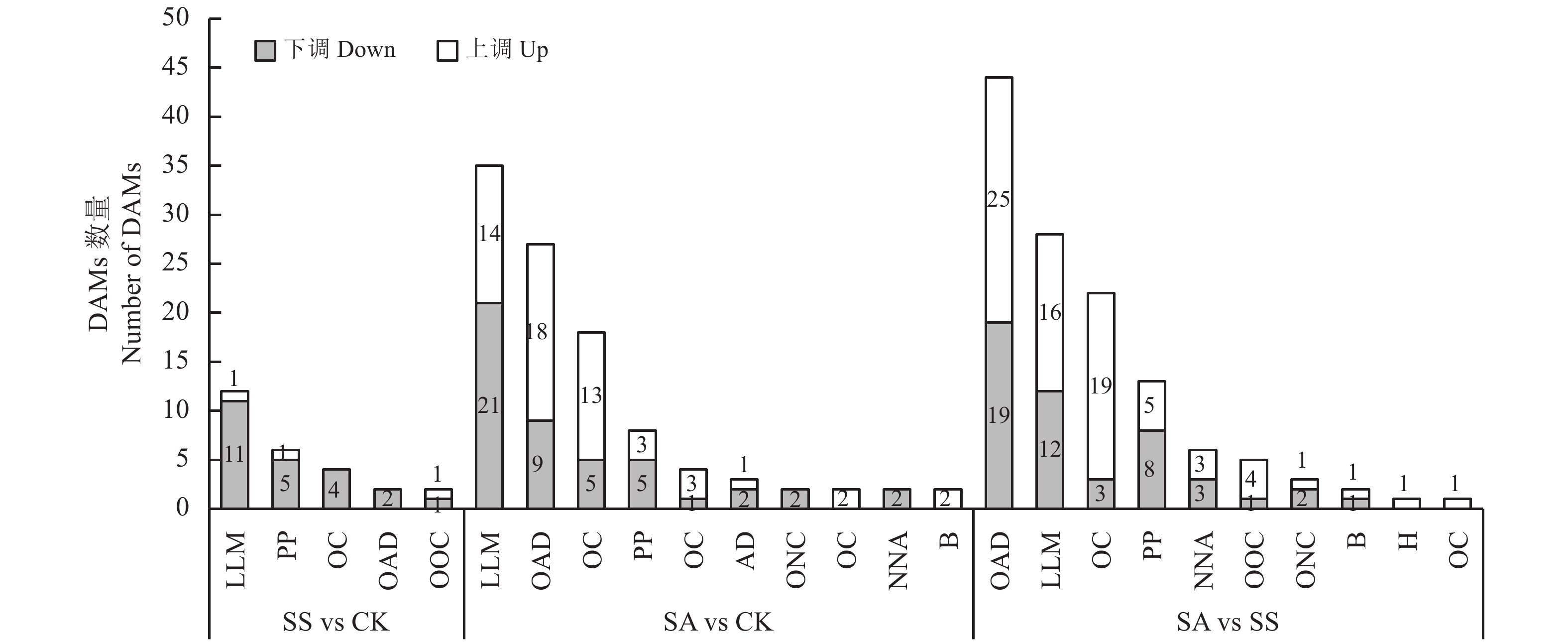
LLM:脂质和类脂分子; PP:苯丙烷类和聚酮类; OC:有机杂环化合物; OAD:有机酸及衍生物; OOC:苯丙烷类和聚酮类;AD:生物碱及其衍生物; ONC:有机氮化合物; NNA:有机杂环化合物; B:有机氧化合物; H:碳氢化合物。
Figure 6. Analysis of differentially accumulated metabolites under salt or alkali treatment
LLM: lipids and lipid-like molecules; PP: phenylpropanoids and polyketides; OC: organoheterocyclic compounds; OAD: organic acids and derivatives; OOC: organic oxygen compounds; AD: alkaloids and derivatives. ONC: organic nitrogen compounds; NNA: nucleosides, nucleotides and analogues; B: benzenoids; H: hydrocarbons.
![]()
图 7 盐、碱胁迫对差异代谢途径的富集结果影响
a:盐胁迫条件下的代谢富集途径;b:碱胁迫的代谢富集途径;c:盐与碱胁迫之间的差异代谢富集途径。富集率是观测差异代谢与期望代谢物之间的比值。
Figure 7. Effect of enrichment results of differential metabolic pathways under salt or alkali stress
a: metabolic enrichment pathway under salt stress; b: metabolic enrichment pathway under alkaline stress; c: differential metabolic enrichment pathways under salt and alkaline stress. The enrichment ratio was computed as Hits/Expected.
-
[1] 杜良宗, 谭昆, 周少梁. 土壤盐碱地改良的研究分析与发展概况. 土壤科学, 2021, 9(1): 41-45. DU L Z, TAN K, ZHOU S L. Research analysis and development of soil saline-alkali land improvement. Hans Journal of Soil Science, 2021, 9(1): 41-45.
[2] 杨劲松. 中国盐渍土研究的发展历程与展望. 土壤学报, 2008(5): 837-845. YANG J S. Development course and prospect of saline soil research in China. Journal of Soil Science, 2008(5): 837-845.
[3] 徐璐, 王志春, 赵长巍, 王明明, 马红媛. 东北地区盐碱土及耕作改良研究进展. 中国农学通报, 2011, 27(27): 23-31. XU L, WANG Z C, ZHAO C W, WANG M M, MA H Y. Research progress of saline-alkali soil and tillage improvement in Northeast China. China Agriculture Bulletin, 2011, 27(27): 23-31.
[4] 刘丽娟, 李小玉. 干旱区土壤盐分积累过程研究进展. 生态学杂志, 2019, 38(3): 891-898. LIU L J, LI X Y. Progress on soil salt accumulation process in arid areas. Journal of Ecology, 2019, 38(3): 891-898.
[5] 燕辉, 彭晓邦, 薛建杰. NaCl 胁迫对花棒叶片光合特性及游离氨基酸代谢的影响. 应用生态学报, 2012, 23(7): 1790-1796. YAN H, PENG X B, XUE J J. Effects of NaCl stress on leaf photosynthesis characteristics and free amino acid metabolism of Heyedysarum scoparium. Journal of Applied Ecology, 2012, 23(7): 1790-1796.
[6] 郭瑞, 周际, 杨帆, 李峰. 碱胁迫对小麦(Triticum aestivum Linn.)叶片代谢过程的影响. 中国农业科学, 2017, 50(2): 250-259. GUO R, ZHOU J, YANG F, LI F. Effect of alkali stress on the leaf metabolic process of wheat (Triticum aestivum Linn.). Agricultural Science of China, 2017, 50(2): 250-259.
[7] 石德成, 盛艳敏, 赵可夫. 复杂盐碱条件对向日葵胁迫作用主导因素的实验确定. 作物学报, 2002, 28(4): 461-467. SHI D C, SHENG Y M, ZHAO K F. Experexperimental determination of the dominant factors of sunflower stress by complex salinity conditions. Journal of Crop Sciences, 2002, 28(4): 461-467.
[8] 杨春武, 李长有, 尹红娟, 鞠淼, 石德成. 小冰麦(Triticum aestivum-Agropyron intermedium)对盐胁迫和碱胁迫的生理响应. 作物学报, 2007, 33(8): 37-49. YANG C W, LI C Y, YIN H J, JU M, SHI D C. Physiological response of small ice wheat (Triticum aestivum-Agropyron intermedium) to salt stress and alkali stress. Journal of Crop Sciences, 2007, 33(8): 37-49.
[9] 麻莹, 曲冰冰, 郭立泉, 石德成, 王德利. 盐碱混合胁迫下抗碱盐生植物碱地肤的生长及其茎叶中溶质积累特点. 草业学报, 2007, 16(4): 25-33. MA Y, QU B B, GUO L Q, SHI D C, WANG D L. Growth of alkali-resistant plants under mixed stress and the characteristics of solute accumulation in stems and leaves. Acta Prataculturae Sinica, 2007, 16(4): 25-33.
[10] DU B, KRUSE J, WINKLER J B. Metabolic responses of date palm (Phoenix dactylifera L.) leaves to drought differ in summer and winter climate. Tree Physiology, 2021, 41(9): 1685-1700. doi: 10.1093/treephys/tpab027
[11] CAI S, WU L, WANG G. DA-6 improves sunflower seed vigor under Al3+ stress by regulating Al3+ balance and ethylene metabolic. Ecotoxicology and Environmental Safety, 2022, 229: 113048. doi: 10.1016/j.ecoenv.2021.113048
[12] ROMHELD V. AWAS F. Significance of root exudates in acquisition of heavy metals from a contaminated calcareous soil by graminaceous species. Journal of Plant Nutrition, 2000, 23: 1875-1866.
[13] ENRIQUE P, LUIS J, CORCUERA. JOSE M. Spatial and temporal variation in citrate and malate exudation and tissue concentration as affected by P stress in roots of white lupin. Plant and Soil, 2002, 241: 209-221. doi: 10.1023/A:1016148222687
[14] 麻莹, 王晓苹, 姜海波, 石德成. 盐碱胁迫下碱地肤体内的有机酸积累及其草酸代谢特点. 草业学报, 2017, 26(7): 158-165. MA Y, WANG X P, JIANG H B, SHI D C. Organic acid accumulation and oxalate metabolism under saline-alkali stress. Acta Prataculturae Sinica, 2017, 26(7): 158-165.
[15] 郭立泉, 陈建欣, 崔景军, 韩丹, 石德成. 盐、碱胁迫下星星草有机酸代谢调节的比较研究. 东北师大学报, 2009, 41(4): 123-128. GUO L Q, CHEN J X, CUI J J, HAN D, SHI D C. Comparative study on the metabolic regulation of star-herb organic acids under salt and alkali stress. Journal of Northeast Normal University, 2009, 41(4): 123-128.
[16] 邹春雷. 盐碱胁迫对甜菜根茎的生理代谢影响. 石河子: 第17届作物生理学术研讨会, 2022. ZOU C L. Effect of saline stress on the physiological metabolism of sugar beet. Shihezi: Abstract of the 17th Crop Physiology Society, 2022.
[17] 戴凌燕, 唐呈瑞, 殷奎德, 阮燕晔, 张立军. 苏打盐碱胁迫对甜高粱植株有机酸含量的影响. 生态学杂志, 2015, 34(3): 681-687. DAI L Y, TANG C R, YIN K D, RUAN Y Y, ZHANG L J. Effect of soda saline stress on organic acid content in sweet sorghum plants. Journal of Ecology, 2015, 34(3): 681-687.
[18] 李俊伟, 刘景辉, 王俊英, 郭来春, 王春龙, 任长忠. 盐与碱胁迫对燕麦离子平衡和有机酸含量的影响. 西北植物学报, 2022, 42(10): 1700-1710. LI J W, LIU J H, WANG J Y, GUO L C, WANG C L, REN C Z. Effect of salt and alkali stress on ionic balance and organic acid content in oat. Journal of Northwest Botany, 2022, 42(10): 1700-1710.
[19] PANG Y Y, YAN H H, GUO L C. Reference genome assemblies reveal the origin and evolution of allohexaploid oat. Nature Genetics, 2022, 54: 1248-1258. doi: 10.1038/s41588-022-01127-7
[20] 景芳, 任生兰, 边芳, 刘彦明, 张成君. 16个燕麦品种在甘肃陇中地区的适应性评价与筛选. 中国草地学报, 2023, 45(1): 68-76. JING F, REN S L, BIAN F, LIU Y M, ZHANG C J. Adaptation evaluation and screening of 16 oat varieties in Longzhong Region, Gansu Province. The Journal of the Chinese Grassland, 2023, 45(1): 68-76.
[21] 王波, 宋凤斌. 燕麦对盐碱胁迫的反应和适应性. 生态环境, 2006, 6(3): 625-629. WANG B, SOMG F B. Oat response and adaptation to saline stress. Ecological nvironment, 2006, 6(3): 625-629.
[22] 杨莉, 赵桂琴, 周向睿, 柴继宽, 杜文盼. 燕麦K+, Na+积累与AsSOS1基因表达对盐胁迫的响应. 草地学报, 2023, 31(2): 337-348. YANG L, ZHAO G Q, ZHOU X R, CHAI J K, DU W P. Oat K+, Na+ accumulation and AsSOS1 gene expression in response to salt stress. Journal of Grassland, 2023, 31(2): 337-348.
[23] 李英丽, 方正, 毛明艳. 不同燕麦品种耐碱性筛选和鉴定. 河北农业大学学报, 2014, 11(6): 17-21. LI Y L, FANG Z, MAO M Y. Basic resistance screening and identification of different oat varieties. Journal of Agricultural University of Hebei, 2014, 11(6): 17-21.
[24] ZHAO Y, LU Z, HE L. Effects of saline-alkaline stress on seed germination and seedling growth of Sorghum bicolor (L.) Moench. Appl Biochem Biotechnol, 2014, 173 (7): 1680-91.
[25] 萨如拉, 刘景辉, 刘伟, 白健慧, 王占海. 燕麦对碱胁迫的阳离子响应机制. 作物学报, 2014, 40(2): 362-368. Sarula, LIU J H, LIU W, BAI J H, WANG Z H. Cationic response mechanism of oat to alkali stress. Journal of Crop Sciences, 2014, 40(2): 362-368.
[26] 刘建新, 刘瑞瑞, 刘秀丽. 盐碱胁迫下外源硫化氢对裸燕麦叶片氨基酸代谢过程的影响. 草业学报, 2023, 32(2): 119-130. LIU J X, LIU R R, LIU X L. Effects of exogenous hydrogen sulfide on amino acid metabolism in naked oat leaves under saline-alkali stress. Acta Prataculturae Sinica, 2023, 32(2): 119-130.
[27] 刘杰, 张美丽, 张义, 石德成. 人工模拟盐、碱环境对向日葵种子萌发及幼苗生长的影响. 作物学报, 2008, 6(10): 1818-1825. doi: 10.3321/j.issn:0496-3490.2008.10.019 LIU J, ZHANG M L, ZHANG Y, SHI D C. The effect of salt and alkali environment on seed germination and seedling growth of sunflower. Crop Journal, 2008, 6(10): 1818-1825. doi: 10.3321/j.issn:0496-3490.2008.10.019
[28] BAI J H, LIU J H, ZHANG N, YANG J H, SA R L, WU L. Effect of alkali stress on soluble sugar, antioxidant enzymes and yield of oat. Journal of Integrative Agriculture, 2013, 12 (8): 1441-1449.
[29] 邹琦. 植物生理学实验指导. 北京: 中国农业出版社, 2003. ZOU Q. Experimental Auidance in Plant Physiology. Beijing: China Agricultural Press, 2003.
[30] 郭家鑫, 鲁晓宇, 陶一凡, 郭慧娟, 闵伟. 棉花在盐碱胁迫下代谢产物及通路的分析. 作物学报, 2022, 48(8): 2100-2114. GUO J X, LU X Y, TAO Y F, GUO H J, MIN W. Analysis of metabolites and pathways in cotton under saline-alkali stress. Journal of Crop Sciences, 2022, 48(8): 2100-2114.
[31] ABBASI, JAMIL, HAQ, ALI, AHMAD, MALIK. Salt stress manifestation on plants, mechanism of salt tolerance and potassium role in alleviating it: a review. Zemdirbyste, 2016, 103(2): 229-238. doi: 10.13080/z-a.2016.103.030
[32] YANG C, CHONG J, LI C, KIM C, SHI D, WANG D. Osmotic adjustment and ion balance traits of an alkali resistant halophyte Kochia sieversiana during adaptation to salt and alkali conditions. Plant Soil, 2007, 294: 263-276. doi: 10.1007/s11104-007-9251-3
[33] FOUGERE F, RUDULIER D L, STREETER J G. Effects of salt stress on amino acid, organic acid, and carbohydrate composition of roots, Bacteroids, and cytosol of alfalfa (Medicago sativa L.). Plant Physiology, 1991, 964: 1228-1236.
[34] BROSCHE M, VINOCUR B, ALATALO E R, LAMMINMAKI A, TEICHMANN T, OTTOW E A, DJILIANWV D, AFIF D, BOGEAT T M B, ALTMAN A, POLLE A, DREVER E, RUDD S, PAULIN L, AUVINEN P, KANGASJARVI J. Gene expression and metabolite profiling of Populus euphratica growing in the Negev desert. Genome Biology, 2005, 6: 681-689.
[35] 王宇超, 王得祥, 彭少兵, 何帆, 余鸽, 王小兰. 盐胁迫对木本滨藜植物细胞膜透性及生理特性的影响. 干旱地区农业研究, 2007, 25(4): 225-229. WANG Y C, WANG D X, PENG S B, HE F, YU G, WANG X L. Effect of salt stress on cell membrane permeability and physiological characteristics of S. benina plants. Agricultural Research in Arid Areas, 2007, 25(4): 225-229.
[36] 栗露露, 殷文超, 牛梅, 孟文静, 张晓星, 童红宁. 油菜素甾醇调控水稻盐胁迫应答的作用研究. 植物学报, 2019(2): 211-219. LI L L, YIN W C, NIU M, MENG W J, ZHANG X X, TONG H X. Role of brassinosterol in regulating salt stress response in rice. Journal of Botany, 2019(2): 211-219.
[37] SCHINDLER M, HAWTHOME F C. A bond-valence approach to the structure, chemistry and paragenesis of hydroxy-hydrated oxysalt minerals. ii. crystal structure and chemical composition of borate minerals. Can Mineral, 2001, 39 (5): 1243-1256.
[38] 王佳敏. 植物中ABA和JA对盐胁迫的响应. 生物过程, 2023, 13(1): 33-38. WANG J M. ABA and JA responses to salt stress in plants. Biological Processes, 2023, 13(1): 33-38.
[39] CORTI E, FALSINI S, SCHIFF S, TANI C, GONNELLI C, ALESSIO P. Saline stress impairs lipid storage mobilization during germination in Eruca sativa. Plants, 2023, 12(2): 366-366. doi: 10.3390/plants12020366
[40] LI Y X, LIU B Y, PENG Y X, LIU C L, ZHANG X Z, ZHANG Z J, LIANG W, MA F W, LI C Y. Exogenous GABA alleviates alkaline stress in Malus hupehensis by regulating the accumulation of organic acids. Scientia Horticulturae, 2020, 261: 108982.
[41] JIA X M, WANG H, SVETLA S, ZHU Y F, HU Y, CHENG L, ZHAO T, WANG Y X. Comparative physiological responses and adaptive strategies of apple Malus halliana to salt, alkali and saline-alkali stress. Scientia Horticulturae, 2019, 245: 154-162. doi: 10.1016/j.scienta.2018.10.017
[42] XING G, MA W, GAO S, JIN Z, YUE Q, YAO Y. Transcriptomic and phosphoproteomic profiling and metabolite analyses reveal the mechanism of NaHCO3 induced organic acid secretion in grapevine roots. BMC Plant Biology, 2019, 19(1): 383. doi: 10.1186/s12870-019-1990-9
[43] 石德成, 盛艳敏, 赵可夫. 复杂盐碱条件对向日葵胁迫作用主导因素的实验确定. 作物学报, 2002, 5(4): 461-467. SHI D C, SHENG Y M, ZHAO K F. Experexperimental determination of the dominant factors of sunflower stress by complex salinity conditions. Journal of Crop Sciences, 2002, 5(4): 461-467.
[44] GHOULAM C, FOURSY A, FARES K. Effects of salt stress on growth, inorganic ions and proline acumulation in relation to osmotic adjustment in five sugar et cultivars. Environmental and Experimental Botany, 2002, 47: 39-50. doi: 10.1016/S0098-8472(01)00109-5
[45] SHORT D C, COLMER T D. Salt tolerance in the halophyte Halosarcia pergranulata subsp. pergranulata. Annals of Botany, 1999, 83: 207-213. doi: 10.1006/anbo.1998.0812
[46] 杨春武, 李长有, 张美丽, 石德成. 盐、碱胁迫下小冰麦体内的pH及离子平衡. 应用生态学报, 2008, 8(5): 1000-1005. YANG C W, LI C Y, ZHANG M L, SHI D C. The pH and ion balance in small ice wheat under salt and alkali stress. Journal of Applied Ecology, 2008, 8(5): 1000-1005.
[47] 焦德志, 赵泽龙. 盐碱胁迫对植物形态和生理生化影响及植物响应的研究进展. 江苏农业科学, 2019, 47(20): 1-4. JIAO D Z, ZHAO Z L. Progress in studying the effects of salinity stress on plant morphology, physiology and biochemistry and plant response. Jiangsu Agricultural Science, 2019, 47(20): 1-4.
[48] YANG C, ZHENG S, HUANG H. Comparison of osmotic adjustment and ion balance strategies in nineteen alkali-tolerant halophyte species during adaptation to salt-alkalinized. Australian Journal of Crop Science, 2012, 6 (1): 141-148.
[49] 高玉刚. 基于转录组和代谢组联合分析燕麦响应盐碱胁迫的机制研究. 大庆: 黑龙江八一农垦大学博士学位论文, 2022. GAO Y G. Study on the responses of ota to saline-alkali stress based on transcriptome and metabolomejoint analysisis. PhD Thesis. Daqing: Heilongjiang Bayi Agricultural ReclamationUniversity, 2022.
[50] 吕家强, 李长有, 杨春武, 胡锐. 天然盐碱土壤对虎尾草茎叶有机酸积累影响及胁迫因子分析. 草业学报, 2015, 24(4): 95-103. LYU J Q, LI C Y, YANG C W, HU R. Effect of natural saline-alkali soil on organic acid accumulation and analysis of stress factors. Acta Prataculturae Sinica, 2015, 24(4): 95-103.
[51] 殷立娟, 祝玲. 羊草苗对盐碱胁迫的反应和适应性. 东北师大学报: 自然科学版, 1989, 5(4): 87-95. YIN L J, ZHU L. Response and adaptability of grass seedlings to saline stress. Journal of Northeast Normal University: Natural Science, 1989, 5(4): 87-95.
[52] 柯梅侯, 钰荣, 张一弓. 碱胁迫下冰草根系pH值与有机酸含量变化. 新疆农业科学, 2021, 58(10): 1929-1937. KE M H, YU R, ZHANG Y G. Changes of pH and organic acid content in ice roots under alkali stress. Xinjiang Agricultural Science, 2021, 58(10): 1929-1937.
[53] 郭瑞, 周际, 杨帆, 李峰, 李昊如, 夏旭, 刘琪. 拔节孕穗期小麦干旱胁迫下生长代谢变化规律. 植物生态学报, 2016, 40(12): 1319-1327. GUO R, ZHOU J, YANG F, LI F, LI H R, XIA X, LIU Q. Changes of growth metabolism under drought stress in wheat during jointing and booting stage. Journal of Plant Ecology, 2016, 40(12): 1319-1327.
-
期刊类型引用(3)
1. 白桦,肖文昌,杨筱筱,张阳,温珍玉,李斌,陈静. 不同数据源对汛期旱涝急转规律的影响及成因——以江西为例. 中国农村水利水电. 2025(03): 30-36 .  百度学术
百度学术
2. 鲍艳磊,吴朝阳,郑东博. 大气污染对京津冀地区夏季植被生长峰值的影响. 地理科学进展. 2023(06): 1124-1138 . 百度学术
3. 侯晓琦. 张家口地区干旱改变特征探析. 农业灾害研究. 2021(06): 110-111+113 . 百度学术
其他类型引用(1)

 下载:
下载:











计量
- PDF下载量: 0
- 文章访问数: 0
- HTML全文浏览量: 0
- 被引次数: 4

