

{kind=link}
{kind=link}
{kind=link}
种子生态学研究现状
[张红香 , 周道玮]
, 周道玮]
, 周道玮]
|
|


第一作者:张红香(1981-),女,辽宁瓦房店人,副研究员,博士,研究方向为种子生态学、草地生态学。E-mail:[email protected]
从种子经历的生命历程——发育、传播、捕食、种子库、休眠与萌发和种子性状6个方面综述种子生态学领域国内外研究现状和一些新结果。对重要的专业术语给出定义及英文注释,涵盖了一些领域内核心的假说、理论和有争议的研究问题,旨在让读者了解种子生态学的主体内容和研究近况。针对研究现状和气候变化背景,建议今后广泛深入开展不同环境和气候条件下母本效应及种子多功能性状间关系的研究。
We summarized overseas and domestic research status and new results in seed ecology, from six aspects including seed traits, seed development, dispersal, predation, seed bank, dormancy and germination, the life processes that seed experienced. The definition and English dictions were given for the important terms. Some central hypothesis, theories and controversial questions were included. We aimed to let the audiences know the main contents and the recent developments in seed ecology. According to the research status and background of climate change, we suggested that maternal effects and the relationship between multiple functional traits of seeds under different environments and climates should be comprehensively and intensively studied.
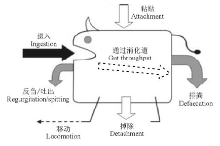
种子生态学是生态学一个年轻的研究分支, 产生于1973年, 标志性事件是1972在英国诺丁汉大学首次召开的种子生态学会议和随后Heydecker编辑出版的《Seed Ecology》论文集[1, 2]。1985年Michael Fenner出版了《种子生态学》专著[3]。种子生态学相似于生态学的定义形式, 表述为研究种子与其环境相互关系的科学[4]。种子是植物体主要的可动器官, 且经历多样化的生命历程, 包括传播、捕食、休眠、萌发、死亡等。为了体现这种特点, 种子生态学可定义为:研究生物与非生物环境影响下的种子生命历程、性状适应与进化的科学。种子从出生到死亡可能经历的过程及时空变迁就是主体研究内容, 包括种子发育、种子传播、种子捕食、种子库、种子休眠与萌发、种子性状适应与进化, 每一个过程都面临着死亡的风险(图1)。非生物因素主要包括气候因素(如温度、降水、光照)和环境胁迫(如盐碱、重金属), 生物因素有动物、植物(包括母株)、微生物及人为干扰(如耕作)。
 | 图1 种子生态学的概念图解Fig.1 Conceptual graph of seed ecology |
种子生态学属于交叉学科, 除了对于生态学、繁殖生物学、进化生物学等学科的学术贡献, 种子生态学研究对于作物种子生产、农业育种、杂草防除、生态恢复、生物多样性保护等方面也具有重要的实践意义。近年来, 在国内外专业学术组织(如国际种子科学学会ISSS和国内植物学会种子科学与技术专业委员会)和专业学术会议(如国际种子生态学大会和全国种子科学与技术研讨会)的推动下, 种子生态学发展较快。
生态学的一个评述性的连续出版物《生态科学进展》第一卷第一个专题即介绍了种子生态学研究进展[5], Fenner和Thompson[6]在1985年Fenner的《种子生态学》基础上, 总结了20年来的新进展, 于2005年再版了《种子生态学》专著。近年来国内外也发表了一些种子生态学领域综述性的文章, 总结了某一类种子的生态学特征(如顽拗性种子)[7]、种子生态学在应用实践中的作用(如杂草管理[8]和恢复生态[9])或种子生态学领域某个重要研究内容的进展(如种子的扩散和传播[10, 11]、土壤种子库[12]、种子休眠与萌发[13, 14]、种子重量生态[15])。黄振英等[1]概述了种子生态学的研究内容、研究途径和发展趋势, 于顺利和方伟伟[16]从种子散布、种子重量、土壤种子库和种子地理学4个研究方向阐述了种子生态学的研究动态。
本文从种子经历的生命过程:发育、传播、捕食、种子库、休眠与萌发5个方面及种子性状综述了种子生态学的研究现状和近几年的一些新研究结果, 对于重要名词术语给出对应的英文词汇, 对一些易混淆或有新内涵的术语加以明确定义, 概述一些领域内的重要假说, 目的是:1)使读者对种子生态学主体内容有综合理解; 2)提供一些新的信息; 3)推动种子生态学在中国的发展。
从种子发育的角度, 种子生态学主要关注种子发育期的外界环境条件对种子性状(如尺寸、鲜干重、活力、休眠性)及子代的影响, 即母本效应(maternal effect)[17]。因种子的扩散能力通常是有限的, 大部分仍分布在母本周围。当母体与子代经历的环境变化呈负相关时即在波动的环境中, 母体会暗示后代改变发育过程或为其提供更多资源以利于存活, 这种母本效应被称为“ 预见性母本效应” (anticipatory maternal effects)或“ 传代表型可塑性” (transgenerational phenotypic plasticity)。当母体与子代经历的环境变化呈正相关时即在相对稳定的环境中, 则会导致预见性母本效应的进化及过去环境的表型记忆(phenotypic memory)。当母体和子代经历的环境变化不相关时, 母本效应的方式是“ 两头下注” (bet-hedging)或“ 风险均摊” (risk-spreading), 产生多样化表型的子代, 被称为“ 随机性母本效应” (randomizing maternal effects)[18]。
母本环境对种子性状的影响代表母本到子代的通路[19], 是生活史的重要一环, 决定着子代的适合度, 因此得到越来越多的关注和研究。研究发现, 种子性状(如休眠性、寿命)与母本在种子成熟及开花结实期间经历的环境条件(如温度和降水)密切相关[20, 21, 22, 23, 24]。母本在种子成熟期经历的温度影响不同光敏色素的表达, 从而影响种子萌发, 因为光敏色素调节种子萌发对光照和温度的响应[20]。母本经历的温度越低, 种子休眠程度越高[6]。对拟南芥(Arabidopsis thaliana)的研究表明, 母本经历的低温能够激活开花基因座位(Flowering Locus T), 抑制果实中原花色素的合成, 使其延迟开花, 从而改变种皮中的单宁含量, 最终引起种子休眠率增加[21]。母本在种子成熟期经历低温引起的休眠, 会造成种子在低水势下发芽能力的降低[22]。然而对西班牙山区一种多年生草本植物Centaurium somedanum的研究发现, 考虑到温度的年际变化, 种子扩散时的休眠程度和对休眠打破因子的敏感性与母本开花期的温度显著正相关[23], 与普遍接受的结论相反。降水对于作物生产非常重要, 但是种子发育和成熟期降水会降低作物种子的活力和寿命[24]。对欧鼠李(Frangula alnus)的研究发现, 开花和结实物候影响随后的种子扩散、萌发和幼苗存活[25]:早开花提高结实率, 晚开花延迟结实但种子更大, 易于传播和种群更新。
传代影响研究的一个重要科学问题是种子和子代的性状差异是母本效应的结果还是环境改变引起的适应响应。有研究发现, 同一物种来源于低地的种子寿命长于来源于高山的, 但是播种到共同的花圃后, 来自高山的二代种子寿命是一代的2倍, 而来自低地的二代种子寿命没有增加。该研究结论是种子寿命有遗传基础, 但可能表现出对环境的强烈适应, 这种响应与亲本效应调控mRNA积累有关[17]。对具有不同生活史的美国风铃草(Campanulastrum americanum)的传代可塑性研究结果表明, 母本和子代的光照条件都影响后代的表型[26]。与母本生长在相同光环境下的子代种子萌发率和幼苗存活率更高。子代种子在光照条件下大多数在秋季萌发, 成为一年生植物, 而在遮阴条件下大多数在春季萌发, 成为二年生植物。Yang等[27]将两种异型性的角果碱蓬(Suaeda corniculata)种子不同时间播种于不同盐度土壤中, 发现晚播种(不利条件)的母体产生的子代繁殖生物量比例高, 产生的不休眠种子更少。与母本播种于相同盐浓度的子代个体比播种于相反浓度的个体繁殖分配高, 种子异型性的传代效应与实验条件相互作用共同决定子代的表型。
种子传播是指种子传播体(diaspore)从母株脱离通过不同媒介传播到不同位点的过程, 包括种子或果实依靠自身的重力或外界风力等散布到地表的过程及外界传播媒介对地表种子搬运的二次迁移过程[28], 又称种子扩散或种子散布[29]。种子传播体主要指种子或包含种子的果实、复合果为主的扩散单元(dispersal unit)。种子传播媒介(vector)包括风、水、动物、人类活动等, 因此种子传播体根据传播表现被归类为风媒(anemochory)、水媒(hydrochory)、动物媒介(zoochory)和自传播种子(autochory)[30]。种子传播研究主要集中于4个方面:种子雨时空格局与影响因素[31, 32, 33, 34, 35, 36], 种子传播模型特别是风传播模型和机制[37, 38, 39, 40, 41], 种子的动物传播及种子传播的进化意义[42, 43, 44, 45, 46, 47]。
在特定的时期内从母株上散落的种子量叫种子雨(seed rain), 通常用种子雨密度这一指标体现, 分为种群种子雨和群落种子雨[31]。种子初始扩散的轨迹分布叫做种子影(seed shadow)[32]。种子雨和种子影的关系相当于量与面积的关系, 在中文文献中, 极少使用种子影, 而在国际上仍使用较多[33]。种子初始扩散受传播体性状、母体性状、周围植被条件、风力条件、湿度、人为干扰等因素影响[32, 35, 36]。这部分内容的研究对象大多集中于木本植物种群或群落。陕西辽东栎(Quercus liaotungensis)群落种子雨密度与物种丰富度显著正相关[34]。胡杨(Populus euphratica)种子雨强度与湿度呈显著负相关, 一天之中, 种子在湿度较低的中午和下午散落[35]。对南美半干旱灌木地种子雨的研究发现, 放牧和火干扰增加种子雨多样性[36]。
种子传播是植物主要的运动机制。过去大部分研究主要集中于短距离传播, 研究其对植物出土定居及物种共存的作用[37]。近年来, 因种子传播对生态和进化时空尺度上(如入侵植物学和集合群落构建过程)的重要性[38], 更多研究转向种子的长距离传播。长距离传播通常指几百米以上的距离, 数据很难获取, 需发展模型解决这一问题。在种子的各种长距离传播方式中, 风对种子的长距离传播是最适合进行模型研究的, 因此, 种子的风传播模型成为传播生态研究的一个重要方向[39]。风传播种子模型通常分为现象模型和机理模型, 现象模型揭示与环境因子交互作用下风的运动与种子传播格局的关系, 而机理模型揭示影响种子传播的每种动力及其机制[40]。最新有研究将地形因素加入机制模型[37], 以及将种子短、中、长距离传播归于统一的模型下[38]。除了研究传播过程的模型以外, 还有模型研究传播体脱离或释放(abscission or release)机制, 因为近来人们意识到传播体脱离或释放决定着后续的传播。研究结果发现, 瞬时风速越高传播体脱离越快, 然而传播体确切的脱离时间还取决于传播体成熟期间经历的风速历史(history of wind speeds)[41]。
动物对种子的传播起重要作用, 除了开拓新位点及扩大种群, 传播种子的动物与种子被传播植物之间还可能存在协同进化关系(co-evolution)[11]。在食果动物与传播植物的相互作用系统中, 中心问题是理解传播效应的功能结果。食果动物的传播效应包括多重效应(multiple)、互补或冗余效应(complementary or redundant)、特异性效应(distinct effects)[42]。研究者们从定性(种子处理、传播、扩散后定居)和定量(动物探访率、采食率)两方面研究了动物传播的效应[43]。但我们对此的理解还很有限[42]。以动物为媒介的传播总是伴随着对种子的捕食, 将在下文详述。人类活动对动物媒种子传播的影响依赖于干扰类型、生物类群及干扰发生的地理区域。一般认为生境破碎化造成食果动物多样性下降[44], 然而Meta分析发现, 生境碎片化和退化对温带地区传播种子的动物多样性具有负面影响, 而对热带地区食果动物多样性没有显著影响, 但是对温带和热带生境的相互作用率(即果实移除率和动物探访率)都有影响[45]。
种子传播的进化意义一直是生态学家和进化学家关注和探索的课题。主要的选择动力包括两方面:减轻家族竞争(Kin competition)与避免近亲交配(inbreeding depression), 占据时空异质性生境[40]。与此对应, 科学家提出3个假说来解释种子扩散的意义, 包括逃逸假说(escape hypothesis)、移居假说(immigration hypothesis)和定向假说(directional dispersal hypothesis)[46]。种子扩散使一些个体逃避母体位点周围的家族竞争, 而去占据由不相关且具有不同扩散习性的个体控制的随机的或特定的位点[47]。因此, 传播与两种代价相关, 扩散者自身的直接代价(如死亡率)和与亲缘个体竞争产生的间接代价[48]。环境的时空异质性会导致种子扩散增加, 以便在不确定条件下两头下注[40, 49], 是随机性母本效应的结果。
捕食是动物主动传播种子的目的, 动物的主动传播形式分为体内(消化道)传播(endozoochory)和贮食传播(synzoochory)[50]。种子的动物传播大部分是捕食后的结果, 被动传播主要依靠动物体的粘附和掉除(图2)[51]。在热带雨林中, 动物捕食是种子死亡的主要原因, 种子扩散后的死亡率通常超过75%[52], 对植物适合度和群落结构有很大影响。从传播动物的种类看, 研究者们更多关注啮齿动物和鸟类对种子的捕食和传播[53, 54, 55, 56, 57]。从内容看, 近来主要研究了动物消化道传播和贮食传播机制及效应[51, 57, 58], 扩散前捕食和扩散后捕食的影响[59, 60, 61, 62, 63, 64, 65], 以及种子或果实大小与捕食动物大小和类型间的关系[66, 67, 68]。
 | 图2 动物传播种子的主要途径模式图[51]。Fig.2 Hypothetical animal illustrating the main ways in which plant propagules are dispersed[51] |
近年来鸟类对种子传播的影响研究受到更多重视, 数据显示鸟类传播物种数以往被大大低估了[53]。鸟类的迁徙能够帮助种子长距离传播, 从而快速扩展物种的分布范围[54]。鸟类对种子的传播还能导致大量外来肉果物种的入侵成功, 因为鸟类主要传播外来肉果类植物到本地肉果类物种之下[55]。在巴拿马中部低地森林中的研究发现, 托姆斯刺鼠(Proechimys semispinosus)和红尾松鼠(Sciurus granatensis)通常是短距离传播种子(< 3 m), 它们更喜欢将种子放到草层茂密的大树下(直径> 10 cm), 果实被吃掉而种子完整。这类小型啮齿动物不是捕食者而更像真正的种子传播者[56]。有些啮齿动物有分散贮食的习性, 为了降低食物因同类密度高而被偷窃的可能性, 会把种子扩散到同类密度较低的位点, 增加种子的存活率[57], 因此增加了种子传播的位点和有效性。
动物体内传播有两个假说, 消化优先模式假说(digestion-priority mode hypothesis)和移动优先模式假说(locomotion-priority mode hypothesis)。前者认为动物移动不会影响种子的排出速率, 后者认为动物移动会延长种子在消化道中保留的时间。基于消化优先模式假说的假设, 绝大部分关于种子在消化道保持和存活率的实验数据都是在休息动物中获取的[51]。然而最近对运动的鲤鱼(common carp)的研究结果支持移动优先假说[58]。
除了传播后捕食(pro-dispersal predation), 传播前捕食(pre-dispersal predation)也占一定比例[59]。动物在种子传播前捕食会影响种子装配(seed packaging)对策, 即大小数量权衡, 进而改变植物的资源分配。对具有两种相反种子装配对策的刺柏(Juniperus thurifera)种群的研究发现, 北非种群球果(果中具有单个大种子)采食率低于南欧种群(果中具有多个小种子)。南欧种群未被捕食的球果中的种子会比被捕食球果中未被吃掉的种子更大且发芽率更高, 说明植物应对扩散前捕食会进化出产生大量小种子的生活史对策[60]。一些多年生植物有大年结实现象(mast seeding or masting)[61], 这种现象也与动物捕食和传播有关。有两种假说来解释这种现象:捕食者满足假说(predator satiation hypothesis)和捕食者传播假说(predator dispersal hypothesis)。前者认为这是植物增加种子传播前存活率的适应对策, 通过满足捕食者而减少种子的被捕食损失比率[62]; 后者认为大年结实增加单位种子传播率及传播距离[63]。两个假说各有证据支持, 争论一直持续到现在。对中国西南亚热带森林油茶树(Camellia oleifera)进行的8年的研究结果支持捕食者满足假说[64], 而对波兰温带森林中的欧洲山毛榉(Fagus sylvatica)进行4年的研究结果则支持捕食者传播假说[65]。
人们普遍认为大种子应该被择优捕食[66], 且大种子易被体型大的动物捕食和传播[67]。然而最近澳大利亚研究者对全球13 135个动物-种子关系综合分析得出:种子大小与捕食动物体重呈负相关。主要原因是有蹄类动物捕食小种子及干种子, 将有蹄类动物或非肉果类果实类型排除, 则关系变为正相关。体型大的动物既捕食大种子, 也捕食较小的种子, 动物体重大小与捕食种子的物种数呈显著正相关关系[68]。
种子库通常指人工种子保存库以外的自然种子库, 包括植冠种子库(canopy seed bank or aerial seed bank)和土壤种子库(soil seed bank)。植冠种子库指植物繁殖体成熟后停留在母体上形成的种子库, 种子可以在植冠上存留1~30年或更长时间, 是火灾易发区和干旱荒漠区植物的主要特征[69]。研究论文中的种子库大多指土壤种子库, 即存在于土壤表面、土壤层中或土壤枯落物中的活种子总和[70], 是植物种群和群落更新的物质基础[12]。土壤种子库通常分为两种类型, 短暂(瞬时)土壤种子库(transient soil seed bank)和持久土壤种子库(persistent soil seed bank)[71]。不同生态系统(森林、草地、农田、荒漠、湿地)中种群、群落种子库及区域种子库的动态、组成和分布研究已经积累了大量数据, 到现在几乎所有植被类型的土壤种子库都被研究过。通常来说, 草地土壤种子库密度大于森林土壤种子库[72], 荒漠或沙地土壤种子库密度较低, 冻原地区土壤种子库密度最低、物种数最少[73]。近几年种子库研究主要集中于以下几方面, 即不同环境(如土壤水分、海拔、地形坡向、林/草地退化)和干扰因素(如土地利用、农作措施、放牧与围封)对土壤种子库的影响[74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89], 土壤种子库与地上植被的关系及在植被恢复中的作用[90, 91, 92], 土壤种子库研究方法的研究和总结[92, 93, 94, 95, 96]。
种子库密度受气候条件如温度的影响, 但不同物种响应不同, 有的物种种子库密度随温差增加而降低, 而对有的物种没有显著影响[74]。沿着干旱梯度研究种子库密度发现, 越湿润位点的土壤种子库密度越高[75], 但是在湿润土壤条件下, 种子活力会由于真菌侵袭而降低[76]。随着海拔增高, 土壤种子库密度和物种丰富度均降低[77, 78]。有研究表明, 长期氮沉降使草地土壤种子库的物种数减少[79], 或种子密度和物种数均减少, 并且停止施氮4年后仍然没有恢复[80]。放牧也会降低草地土壤种子库密度[81, 82], 但是不排除一些情况下, 中等放牧增加土壤种子库密度[83]。耕种对森林二次演替过程中土壤种子库影响的研究发现, 土壤种子库密度和物种丰富度随着休耕年限增加而增加, 耕作周期和耕地方式影响不大[84]。中国三江平原地区湿地开垦为农田后, 耕种时间与土壤种子库密度和物种丰富度呈反比例关系[85]。除了管理方式对种子库有显著影响外, 土壤特征也影响种子库组成和多样性。对松嫩草地羊草(Leymus chinensis)优势群落种子库的研究发现, 碱度和Na离子含量与种子库密度和物种丰富度显著负相关[86]。关于严重的干扰事件后土壤种子库的物种组成变化有两个假说, 第一个假说是生态叠加假说(ecological palimpsest hypothesis), 认为种子库的物种丰度会逐渐增加。草地放牧和割草后的恢复研究结果符合生态叠加假说[87, 88]。第二个假说是群落分解假说(community disassembly hypothesis), 认为种子库的物种会逐渐损失, 种子出土用于地上群落构建。森林砍伐后恢复为半天然草地的过程中, 种子库经历砍伐干扰后的调查结果支持群落分解假说[89]。
生态和进化学家认为土壤种子库是生物多样性库(biodiversity reservoirs), 对于种群维持和群落动态有重要意义[90]。相反, 保护和恢复生态学的研究经常发现种子库有微弱的保存价值或恢复潜力[91]。对于这样的结果, 有研究者认为是由于种子库与植物群落取样的系统性偏差造成的, 用物种-面积关系作为工具评估和比较土壤种子库与植物群落单位面积物种丰度发现, 种子库的物种多样性高于地上植被[90], 肯定了种子库对于种群动态和多样性维持的重要作用[92]。
研究方法本身就是重要的研究内容[93]。文献中土壤种子库的取样大小和数量、取样时间、鉴别方法都不一致, 是限制全球种子库特征整合分析的主要因素。取样样方大小有1 m× 1 m、100 cm× 50 cm、50 cm× 50 cm、25 cm× 25 cm、20 cm× 20 cm、10 cm× 10 cm, 还有土芯法取样, 土芯直径1.9~8.0 cm。样方和土芯取样数量在5~60个不等。通常遵循大数量小样方、小数量大样方或大样方内再取小样方的方法。有研究发现, 对于森林土壤种子库最低取样面积为4 m2, 且小数量大样方比大数量小样方更有效[94]。由于种子雨和种子萌发有季节动态, 取样时间不同, 种子库的密度和物种丰富度都不同。很多种子库研究是一年一次或两次取样的结果, 在当年种子雨散布后(短暂与持久种子库之和)或(和)种子开始萌发前取样[72]。一年多次取样和多年长期取样应成为未来方向, 因为土壤种子库季节动态研究和长期定位研究对于揭示种群更新和群落动态具有重要意义。土壤种子库的鉴别方法最常用的是种子萌发法, 国内外研究中90%以上采用萌发法鉴别种子库[95]。然而, 不同物种的休眠类型和萌发条件不同, 得出的种子种类和数量较实际低[96], 应采用物理分离加镜检法、四唑氟化物染色、化学试剂萃取等方法加以辅助鉴别[93], 以求更真实地反映实际的种子库组成和密度。未来需要更多关于土壤种子库研究方法的研究, 得出针对不同生态系统的合理的取样和鉴定标准, 以便区域或全球尺度上分析种子库的特征。
在一定的时间内, 具有生活力的完整种子在水分、温度、光照和氧气等适宜的环境条件下不能萌发的现象叫休眠[97]。种子萌发是指种子吸水开始, 到胚根伸出种皮的过程[98]。萌发完成时幼苗生长才开始, 种子生理学家和生态学家通常只用这个萌发的定义。我国和国际种子检验规程规定当种子发育长成具备正常主要构造的幼苗才称为发芽[99, 100], 原因是种子技术工作者主要关注幼苗的定居能力。对于一粒种子, 休眠和萌发是一种非此即彼的关系, 或称全或无的事件[14]。对于一个种子批(seed lot)或种子群(seed population)而言, 则体现为萌发率(germination percentage)和休眠率。种子休眠能够确保物种在恶劣的环境中存活, 减少同一物种中个体之间的竞争, 以及防止种子在不适宜幼苗生长和定居的时间和地点萌发[101], 是植物的一种生活史对策。但是对于农业生产, 种子收获后休眠就是需要解决的一大障碍。大量的种子休眠研究集中于单个物种或几个物种的休眠原因、类型及破除方法[102, 103], 不同地域和不同物种休眠研究数据的积累已经能够对种子休眠地理和进化进行探讨[104], 种子休眠的季节动态和激素调控已经深入到分子水平[105, 106]。
Baskin 和Baskin[97]将种子休眠分为5 种类型, 包括生理休眠(physiological dormancy, PD)、形态休眠(morphological dormancy, MD)、形态生理休眠(morphophysiological dormancy, MPD)、物理休眠(physical dormancy, PY)和复合休眠(physical plus physiological dormancy, PY+PD), 其中各种休眠类型还被细分为不同的亚类和水平。这是目前广泛认可和应用的种子休眠分类方式。种子随着外界环境的季节性变化, 调整休眠状态, 以选择在有利于幼苗存活的最适宜时间萌发, 表现为休眠的循环, 这种对策也维持了持久土壤种子库[107]。研究发现, 赤霉素GA 通过与脱落酸ABA 及环境因子的复杂作用调节休眠的释放和种子萌发[101]。对模式植物拟南芥的研究表明, 休眠深度与基因表达方式和土壤温度的季节变化有关。冬季土壤温度降低, ABA合成和GA分解代谢基因表达增加, 休眠增加, 种子进入深度休眠。春季和夏季, 土壤温度增加, ABA分解和GA合成基因增加, 休眠解除, 种子萌发[108]。外界的碳和氮水平也可以通过调节激素合成与分解来影响种子的休眠与萌发[109]。
Baskin和Baskin[97]对已发表数据的13 634种植物的全球尺度整合分析发现, 平均70.1% 的植物种子休眠, 29.9%的物种不休眠。热带雨林区不休眠物种比例最高, 为48.7%。随着温度和降水量的降低, 休眠比例增加, 不休眠比例降低, 冷荒漠区只有5%的物种种子不休眠。从休眠类型看, PD是最重要的一类, 特别是在荒漠区和草原区。热带萨瓦纳地区物理休眠的物种比例最高。其它3种休眠类型(MD, MPD, PY+PD)在各种植被类型中几乎都存在, 但是比例很低(图3)[104]。研究揭示, MPD可能是最古老的休眠状态, PD是进化的中心, 其它休眠类型与PD可以相互转化。不休眠是最近的进化发展或者是暂时的状态[110]。
 | 图3 不同类型的休眠种子和不休眠种子的全球生物地理分布[104]Fig.3 World biogeography of morphological, morphophysiological, physical, physical+physiological and physiological dormancy and of nondormancy in seeds[104] |
种子萌发是种子生态学的核心内容, 主要由于它是连接种子和成株的关键一环。种子萌发研究的主要内容是不同外界因素对种子萌发的影响, 包括温度、光照、土壤水分、盐碱胁迫、层积处理、引发处理、贮藏时间和条件、重金属、土壤结皮、射线辐照、激素处理、化学物质处理、微生物制剂、磁场作用等, 在这些因素中, 温度、水分、火对萌发的影响关注度最高。植物种子总是努力选择最合适的条件(包括时间和空间)萌发, 使得种群最大化。作物种子合适的萌发时间对于农业生产也至关重要, 收获前萌发会造成减产。因此, 关键科学问题是种子萌发时间是如何受环境因子调控的。
温度是影响种子萌发时间的最重要的环境因子, 这种影响是复杂的, 因为种子成熟时经历的温度(母体环境温度)和扩散后的温度共同决定着种子的萌发时间[111]。通常母体环境温度高, 则萌发率高[112], 物种生境温度还与萌发最低温(base temperature, Tb)呈显著负相关[113]。植物的季节性萌发也主要与温度有关, 研究发现, 温带地区植物选择春季萌发, 主要是因为春季每日增加的温度促进萌发[114]。季节性突发的热浪(heat wave)对种子萌发也有很大影响, 如果没有热浪事件, 意大利高山植物主要在春季萌发。经历秋季突发热浪能够增加来年春季萌发率, 并且引发植物种子秋季的萌发[115]。
对于扩散后温度对种子萌发的影响, 研究者们提出了种子萌发的积温模型(thermal time model)[116], 模型中最重要的参数是种子萌发最低温(base temperature, Tb)、最适温度(optimal temperature, To)和最高温(ceiling temperature, Tc)及积温(thermal time constant, θ )。植物的积温特点受它们地理起源的影响, 热带物种通常积温低于温带物种, 发芽最低温高于温带物种, 因此热带物种比温带物种发芽快[117]。青藏高原12种菊科植物的发芽最低温较低、积温较高, 也是长期适应青藏高原特殊的温度环境的结果[118]。另外, 发芽需要的温度可能与系统发生有关。禾本科和莎草科植物分别具有较低和较高的最低发芽温度[119]。6种豆科三叶草的发芽积温低于4种禾草羊茅(Festuca ovina)、紫羊茅(Festuca rubra)、直立雀麦(Bromus erectus)和芨芨草(Achnatherum calamagrostis)[120]。C4植物的发芽最低温明显高于C3植物[121]。储藏时间长的种子最低温和最适温度增加[122]。近年来有研究者利用室内培养法结合积温模型模拟和预测全球增温对植物种子萌发和种群更新的影响[123, 124, 125], 这一方法不失为气候变化研究的重要和独特视角。
水分是植物种子萌发的先决条件, 降水量是物种地理分布的主要驱动力之一, 物种的地理分布模式与种子萌发和幼苗出土期对环境(如干旱)的耐受性有关[126]。研究者随积温模型之后提出水势模型(hydrotime model)[127]并组合成水热模型(hydrothermal time model)来刻画种子对温度、水分及其交互作用的萌发响应。Kö chy 和Tiebö rger[128]用水热模型分析地中海地区36 种一年生植物最低温和最低水势的差别, 黄文达等[129]用水热模型分析了阿拉善地区3种荒漠植物的水热响应。水热模型还被用来研究生理休眠种子的休眠释放过程[130]及具有物理休眠种子对温度、水势的萌发响应[131]。Dü rr等[132]从文献和未发表数据中收集了全球243个物种的萌发数据包括水热模型参数, 发现除了模型参数与物种的地理气候起源有关外, 物种萌发速度与温度阈值负相关, 温度与水势阈值负相关。由于模型的假设和分析方法等缺陷, 这3个模型一直受到质疑, 研究者们提出高斯分布、S形温度模型、非线性回归模型、空间网格化、修正积温模型等新模型[133, 134, 135, 136], 都没有得到广泛应用。模型本身及不同物种模型参数的研究热度刚过, 科学家们已经开始将植物的模型参数与其在野外的分布和种群动态数据联系起来, 由推测走向实验验证。最近对荒漠一年生植物的研究表明, 物种的萌发最低温、最适温、最高温、积温、最低水势与其野外20年平均发芽百分数、25年平均发芽日期、种子大小及植株出生死亡统计变化强烈相关, 证实种子的萌发特征对荒漠植物种群动态具有重要的贡献[137]。对巴塔哥尼亚3种山毛榉属(Nothofagus)植物的研究表明, 种子对温度的休眠和萌发响应模式及积温特点决定着它们在野外不同海拔的分布及季节性出土动态[138]。
火对种子萌发的影响在非洲和南美热带稀树草原和森林等生态系统中被广泛研究, 以往的大部分研究结果是温度增加和烟能够打破休眠、促进种子萌发[139, 140]。而对于马达加斯加高地硬叶林的大部分物种, 其种子萌发受到高温和干热(dry heat)的抑制[141]。对历史以来火灾频发的巴西热带高草草原物种的研究发现, 火引起的高温和烟并不能促进植物种子的萌发, 然而种子却并没有被高温杀死, 而是发展出一种火耐受对策[142]。对新热带区易发生火灾的草地生态系统的研究发现, 不同科的优势草本植物采取不同的休眠和萌发对策。有的植物种子不休眠且快速萌发, 有的植物种子萌发率低且有高比例的无活力胚或无胚种子存在, 有的植物种子烟能促进其萌发, 研究首次发现了新热带区植物种子具有生理休眠[143]。阿根廷Chaco季节性干燥森林的26种优势植物对于火干扰也发展出热耐受型萌发(17种)、热促进型萌发(7种)和热敏感型萌发(2种)对策[144]。
种子萌发方面的信息不仅具有学术意义, 还有经济和金融价值。了解什么因素控制萌发时间可以加强1)有效控制杂草的计划性, 2)自然界有重要经济价值的树林、灌木、藤本植物、非禾本草本植物和草本植物的繁殖和传播, 3)受损生态系统的改良[98]。研究发现, 种子起源影响萌发和幼苗存活, 因此, 为了避免不适应环境条件的风险, 受损生态系统恢复最好使用乡土植物的种子[145]。
种子性状包括种子的尺寸、形状、表面及其附属物、颜色、种皮厚度、储藏物质等, 不仅表达了一定量较为稳定的遗传信息, 而且与种子的生产、脱落、传播、储备、萌发等过程密切相关。这些特征反映植物所处的环境条件和适应能力, 决定植物幼苗定居、种群动态, 乃至群落组成和生态系统功能。种子性状中对种子重量的研究最多, 包括环境因素纬度、坡向、海拔和土壤pH 对其产生的影响[146, 147, 148, 149, 150, 151]。大多数研究者主要关注种子单一性状对重要生态过程的影响或相互关系[152, 153, 154, 155], 很少把种子形态、种子传播、萌发对策等多性状有机结合起来进行研究[156]。种子粘液(seed mucilage)[157, 158, 159, 160, 161, 162]、种子异型性(seed heteromorphism)[163, 164, 165, 166, 167, 168]和种子库持久性与种子大小的关系[169, 170, 171, 172, 173]三方面内容是种子性状领域中很好的研究例证。
种子粘液是由种子或果实产生的果胶类物质, 在至少230个属的植物中有过报道。种子粘液对于种子成熟、扩散、种子库维持、休眠、萌发和幼苗生长都有重要作用[157]。最新进展主要是对于这些作用机理方面的揭示。种子粘液使得种子对捕食动物的吸引力降低, 使种子黏附在土壤上难以移动, 并且由于湿滑使种子容易在捕食动物运输过程中掉落[158]。种子粘液粘附在土壤上, 增加土壤持水力和流变性, 有利于维持土壤种子库[159]。种子粘液作为物理屏障, 通过调节进入种子内部的水分和氧气, 避免种子在低温、干旱、高盐等不适宜的条件下萌发[160]。种子粘液能够被土壤中的微生物分解成CO2和可溶性糖, 被种子和幼苗吸收利用, 增加荒漠植物在干旱和沙埋条件下的幼苗出土和早期生长, 降低幼苗死亡率[161, 162]。
种子异型性指同一植株产生不同形状、结构、颜色、大小, 具有不同的传播、休眠、萌发和幼苗出土等行为的种子的现象, 已经在近300个物种中有报道[163, 164]。通常荒漠植物或盐生植物具有两型性种子。如荒漠一年生植物异果芥(Diptychocarpus strictus), 产生一种在植株上部裂开的长角果, 果皮薄、种子翼大、粘液层厚、扩散距离远、萌发率高; 另一种下层不开裂果实, 果皮厚、种子无翼、粘液层薄、扩散距离近、萌发率低[165]。冷荒漠盐生植物角果碱蓬(Suaeda corniculata)的异型性种子通常一种种子成熟时不休眠, 形成短暂种子库, 主要用于幼苗更新, 盐和旱降低其种子活力; 另一种种子具有年际的休眠/不休眠循环, 形成持久种子库, 用于种群维持, 盐和旱会导致其休眠[166]。一年生盐生植物海蓬子柳(Salicornia ramosissima)也具有异型性种子, 花序中部大种子和花序侧面小种子。在低盐条件下, 两种异型性种子发芽率差异不显著, 而在高盐条件下, 中部大种子发芽率显著高于侧面小种子[167]。盐地碱蓬(Suaeda salsa)具有棕色种子和黑色种子两种异型性种子, 较高的GA含量和较低的ABA敏感性, 使得棕色种子在盐胁迫条件下的萌发率高于黑色种子[168]。
种子库持久性与种子大小是否相关一直没有确定的结论[169], 大部分研究支持小种子在土壤种子库中更持久[155, 170], 但有的研究认为大种子更持久或种子大小与寿命和种子库持久性没有显著关系[171]。近来有研究发现, 种子寿命与种子内含物的大小比例有关, 即胚大胚乳小的种子寿命更长[172]。增加种皮厚度指标能够提高种子大小对种子库持久性的预测力, 研究结果表明, 种皮厚的小种子比种皮薄的大种子休眠性强、在种子库中更持久[173]。这些结果都说明, 需要对种子多性状关系进行广泛深入的研究, 综合分析种子的命运及其在种群、群落和生态系统中的作用与使命。
在种子生态学中, 种子库及种子休眠与萌发研究已经积累了大量数据, 数据整合分析以得出大尺度的结果和系统发生学解释对于种群、群落和生态系统生态学和进化生物学都将有重要贡献。种子传播和捕食还缺乏不同地域、不同植物和动物种群、群落的实验研究, 对于传播模型、传播和捕食机理的研究也应加强。在全球气候变化的背景下, 不同气候和环境条件下母本效应、传代效应及种子多性状关系的研究需要更多关注, 以便更好地理解植物的适应与进化, 加强我们对气候变化引起的植被和生态系统改变的预测力。
本文只是综述了种子生态学研究的冰山一角, 或许我们已经进行的研究和得出的结论, 也只是揭示了种子命运及种子生态学理论和实践的冰山一角, 还有很多未解之谜等着我们研究和探索。期待更多的研究生和科技工作者加入种子生态学领域, 通过加强这一分支和交叉学科的研究, 更好地理解和推动群落生态学、恢复生态学、入侵生态学、保护生物学和进化生物学等学科的发展。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [98] |
|
| [99] |
|
| [100] |
|
| [101] |
|
| [102] |
|
| [103] |
|
| [104] |
|
| [105] |
|
| [106] |
|
| [107] |
|
| [108] |
|
| [109] |
|
| [110] |
|
| [111] |
|
| [112] |
|
| [113] |
|
| [114] |
|
| [115] |
|
| [116] |
|
| [117] |
|
| [118] |
|
| [119] |
|
| [120] |
|
| [121] |
|
| [122] |
|
| [123] |
|
| [124] |
|
| [125] |
|
| [126] |
|
| [127] |
|
| [128] |
|
| [129] |
|
| [130] |
|
| [131] |
|
| [132] |
|
| [133] |
|
| [134] |
|
| [135] |
|
| [136] |
|
| [137] |
|
| [138] |
|
| [139] |
|
| [140] |
|
| [141] |
|
| [142] |
|
| [143] |
|
| [144] |
|
| [145] |
|
| [146] |
|
| [147] |
|
| [148] |
|
| [149] |
|
| [150] |
|
| [151] |
|
| [152] |
|
| [153] |
|
| [154] |
|
| [155] |
|
| [156] |
|
| [157] |
|
| [158] |
|
| [159] |
|
| [160] |
|
| [161] |
|
| [162] |
|
| [163] |
|
| [164] |
|
| [165] |
|
| [166] |
|
| [167] |
|
| [168] |
|
| [169] |
|
| [170] |
|
| [171] |
|
| [172] |
|
| [173] |
|