

{kind=link}
{kind=link}
柱花草EST-SSR标记在山蚂蝗中的可转移性与应用
[罗小燕 , 张瑞花, 贾庆麟, 王文强, 白昌军, 丁西朋]
, 张瑞花, 贾庆麟, 王文强, 白昌军, 丁西朋]
, 张瑞花, 贾庆麟, 王文强, 白昌军, 丁西朋]
|
|


第一作者:罗小燕(1980-),女,湖北罗田人,研究实习员,硕士,主要从事热带牧草种质资源评价与选育。E-mail:[email protected]
为鉴定山蚂蝗( Desmodium)种质资源的遗传背景及亲缘关系,选取92对柱花草( Stylosanthes spp.)EST-SSR标记和来自8个种的16份山蚂蝗材料,分析柱花草EST-SSR标记在山蚂蝗中的通用性,并对8种山蚂蝗进行亲缘关系分析。研究结果表明,柱花草EST-SSR标记在8种山蚂蝗种的可转移率为63.04%~73.91%,其中50对可在全部8种山蚂蝗中进行有效扩增。16对多态性EST-SSR标记在16份山蚂蝗种质中共检测到等位基因35个,平均2.19个。聚类分析表明,16份山蚂蝗种质可分为5类,与形态学划分结果相似。可见,柱花草EST-SSR标记在山蚂蝗上具有较高的可转移性,可应用于山蚂蝗种质资源评价及亲缘关系研究。
To identify genetic background and relationships of Desmodium varieties, 92 Stylosanthes EST-SSR markers and 16 varieties from 8 Desmodium species were used to assess the transferability of Stylosanthes EST-SSR markers in Desmodium genus and analyze the genetic relationships among 8 Desmodium species. The results indicated that the transferability of Stylosanthes EST-SSR markers across 8 Desmodium species ranged from 63.04% to 73.91%, and 50 markers of them could produce reliable amplification in all 8 Desmodium species. A total of 35 alleles were generated by 16 polymorphic markers with an average of 2.19 among 16 Desmodium varieties. Cluster analysis showed that 16 Desmodium varieties can be divided into 5 clusters, which was similar to the classification based on morphology. Therefore, Stylosanthes EST-SSR markers showed high transferability in Desmodium, and it would be suitable for germplasm evaluation and genetic relationships analysis of Desmodium.
山蚂蝗属(Desmodium)植物为一年或多年生草本或灌木豆科植物, 起源于东亚、墨西哥和巴西, 是热带亚热带地区重要的植物资源[1]。山蚂蝗具有适应强、生长迅速、生物量大、叶片蛋白含量高的特点, 是极具潜力的高蛋白饲料资源, 具有较好的饲用和综合利用价值[2, 3]。山蚂蝗属植物在全球约有350种, 而在中国分布有27个种5个变种[4], 多数山蚂蝗为二倍体(2n=2x=22)[5]。目前, 关于山蚂蝗植物系统发育研究主要以形态学分类为主, 但山蚂蝗植物种类繁多, 有些种和亚种间的形态特征非常相近, 形态学鉴定非常困难, 存在较多同种异名或同名异种的现象[6, 7]。国内外有学者利用随机扩增多态性DNA(random amplified polymorphic DNA, RAPD)、扩增片段长度多态性(amplified fragment length polymorphism, AFLP)、扩增共有序列遗传标记(amplified consensus genetic markers, ACGM)及同工酶等分子标记对山蚂蝗种质资源进行了遗传多样性及种间亲缘关系分析, 但因研究手段和材料的不同, 其研究结果也存在一定差异, 所以开展研究山蚂蝗属种间的亲缘关系对山蚂蝗种质资源研究与利用尤为重要[8, 9, 10, 11]。
简单重复序列(simple sequence repeat, SSR), 又称微卫星(microsatellite), 是一类由1~6个碱基组成的串联重复序列, 基于SSR序列的SSR标记, 是一种基于DNA长度多态性的分子标记技术[12]。根据其来源的不同, SSR标记可分为基因组SSR标记和EST-SSR标记两种[13]。SSR标记具有数量丰富、多态性高、共显性、操作简单等多个优点, 是种质资源遗传多样性分析、遗传连锁图谱构建、基因定位和分子标记辅助育种的理想工具[13, 14, 15, 16]。但SSR标记的开发成本较高, 在遗传背景研究较薄弱的物种中应用非常有限。分子标记的种间通用性是降低其开发成本, 提高利用效率, 促进遗传学基础较薄弱物种分子遗传学研究的重要途径[17]。山蚂蝗和柱花草(Stylosanthes spp.)作为我国南方重要牧草资源, 均为豆科蝶形花亚科的草本或灌木植物, 亲缘关系较近。因此, 本研究以前期开发的柱花草EST-SSR标记为基础[18], 通过属间通用性分析开发在山蚂蝗中可用的SSR标记, 并对山蚂蝗种间亲缘关系进行分析, 本研究将为山蚂蝗遗传学研究提供重要的分子标记工具, 为其系统发育学研究提供更多分子水平上的依据。
本研究所用材料为来自8个种的16份山蚂蝗材料(表1), 其中7份为卵叶山蚂蝗, 2份圆叶舞草, 2份绒毛山蚂蝗, 异果山绿豆、异叶山蚂蝗、度尼山蚂蝗、糙伏山蚂蝗和灰色山蚂蝗各1份。全部材料均由中国热带农业科学院热带作物品种资源研究所热带牧草研究中心提供。本研究选用的92对柱花草EST-SSR引物均由上海生工生物工程技术服务有限公司合成(表2), 试验中所用的dNTPs、Taq DNA聚合酶、10× Buffer(含Mg2+)及Marker DL 2000等试剂均购自北京全式金生物技术有限公司。
| 表1 16份供试山蚂蝗材料信息 Table 1 Information of 16 Desmodium materials in this study |
| 表2 92对柱花草EST-SSR标记信息 Table 2 Information of 92 EST-SSR markers from Stylosanthes |
从试验基地选择新鲜的山蚂蝗幼嫩叶片, 迅速放入冰盒带回实验室, 利用改良CTAB法进行山蚂蝗基因组DNA抽提[19]。通过1.0%的琼脂糖凝胶电泳和NanoVue超微量分光光度计(GE healthcare)对基因组DNA进行质量和浓度测定, 然后将浓度统一稀释至100 ng· μ L-1, 置于-20 ℃保存备用。
PCR扩增体系和反应程序均参照丁西朋等[20]所述。扩增体系为20 μ L, 包括:100 ng· μ L-1的基因组DNA 1 μ L, 5 μ mol· L-1的正向和反向引物各1 μ L, 10× PCR Buffer(含Mg2+)2 μ L, 10 mmol· L-1的dNTPs 0.4 μ L, 5 U· μ L-1的Taq聚合酶0.3 μ L, 剩余体积用ddH2O补足。反应程序为:94 ℃预变性5 min; 94 ℃变性30 s, 55 ℃退火30 s, 72 ℃延伸40 s, 38个循环; 72 ℃延伸5 min, 4 ℃保存。PCR扩增产物经8%聚丙烯酰胺凝胶电泳后, 通过银染法染色, 拍照记录后统计数据。
用Marker DL 2000作为扩增产物分子量的参照, 采用“ 0, 1” 系统对清晰的电泳条带进行统计, 在同一位点处, 没有条带的记为“ 0” , 有条带的记为“ 1” 。将统计结果汇总到Excel表格中建立二进制矩阵。利用POPGENE 1.32软件进行分析, 计算每对多态性标记的遗传多样性参数, 包括等位基因数(number of alleles, Na)、等位基因频率(allele frequency)、观察杂合度(observed heterozygosity, Ho)、期望杂合度(excepted heterozygosity, He)和Shannon指数(I)[21]。根据等位基因频率按公式(1)计算每个标记的多态性信息含量(polymorphic information content, PIC)。
PIC=
式中:n表示每个位点检测到的等位基因数量, Pi、Pj表示第i、j个等位基因出现的频率[22]。利用NTSYS-pc2.1e软件, 分析材料间的遗传相似系数(genetic similarity, GS), 然后利用GS采用非加权数据分析法(UPGMA)对供试材料进行聚类分析[23], 并通过FreeTree软件[24]采用1 000次重抽样对聚类图进行bootstrap分析。
选用92对柱花草EST-SSR标记对16份山蚂蝗材料进行PCR扩增(图1), 结果表明, 92对标记中有70对标记在山蚂蝗中获得了条带清晰且大小合适的条带, 确定为有效引物, 占总标记的76.1%。剩余22对标记中有7对有多条带(RM40、RM118、RM215、RM377、RM449、RM478和RM491)、15对无扩增产物。
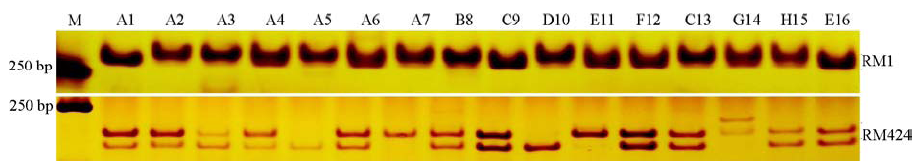 | 图1 部分EST-SSR标记聚丙烯酰氨凝胶电泳结果Fig.1 Polyacrylamide gel electrophoresis of partially SSR markers |
不同柱花草EST-SSR标记在山蚂蝗属中的可转移率有所差异。在70对有效引物中, 有20对在至少1种山蚂蝗中无扩增产物(表3), 在山蚂蝗属中可转移率为37.5%~87.5%。其余50对标记(占总标记的54.35%)在8种山蚂蝗中均可检测到, 表明柱花草EST-SSR标记在山蚂蝗属中具有较高的通用性。而92对柱花草EST-SSR标记在不同种山蚂蝗间的可转移性同样存在明显差异, 在8种山蚂蝗中的可转移率为63.04%~73.91%。由此可知, 不同种山蚂蝗的遗传背景存在显著差别, 因而检测到柱花草EST-SSR标记的数量也不同, 这表明筛选出的可转移性柱花草EST-SSR标记可以对不同种山蚂蝗进行有效区分。
| 表3 20对柱花草EST-SSR标记在8种山蚂蝗间的可转移性 Table 3 Transferability of 20 EST-SSR markers from Stylosanthes across 8 Desmoduim species |
在70对有效标记中共检测到16对标记在16份山蚂蝗材料间具有多态性(表4)。16对多态性SSR标记在16份山蚂蝗材料中共检测到等位基因35个, 平均每个标记2.188个。各多态性标记的观察杂合度(Ho)和期望杂合度(He)分别为0.000~0.813和0.254~0.647, 平均值分别为0.464和0.464。各多态性标记的PIC值为0.215~0.551, 平均值为0.360, 其中RM243的PIC值最高。各多态性的Shannon指数变化范围在0.410~1.036, 平均值为0.672, 同样在RM243中存在最大值, 表明标记RM243具有较高的多态性。
| 表4 16对多态性EST-SSR标记遗传参数 Table 4 Parameters for 16 polymorphic EST-SSR makers |
利用NTSYS-pc 2.10e软件对山蚂蝗种质进行遗传相似系数分析表明, 16份山蚂蝗种质间的遗传相似系数为0.523~0.952, 平均0.711。其中绒毛山蚂蝗E11与E16之间遗传相似性系数最大, 卵叶山蚂蝗A6和异叶山蚂蝗D10之间遗传相似性系数最小。对每份山蚂蝗材料与其它材料之间的平均遗传相似系数分析发现, 度尼山蚂蝗F12与其它材料的平均遗传相似系数最小, 为0.621, 说明度尼山蚂蝗F12与其它7种山蚂蝗材料的亲缘关系较远。基于EST-SSR标记数据的UPGMA聚类分析表明, 在相似系数为0.730时, 可将16份山蚂蝗属材料明显划分为五大类(图2):A类包含7份卵叶山蚂蝗和2份圆叶舞草; B类包含1份糙伏山蚂蝗和1份灰色山蚂蝗; C类由2份绒毛山蚂蝗材料组成; D类包括1份异果山绿豆和1份异叶山蚂蝗; E类只含有1份度尼山蚂蝗。
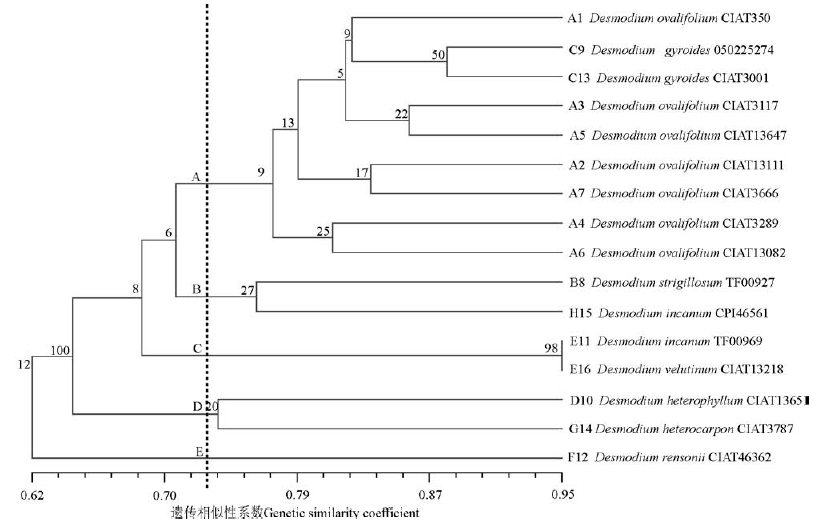 | 图2 基于EST-SSR标记的16份山蚂蝗种质的UPGMA树状图Fig.2 Dendrogram of 16 Desmodium germplasm based on EST-SSR analysis |
利用EST-SSR数据矩阵对这8种山蚂蝗进行相似系数和遗传距离分析发现(表5), 8种山蚂蝗间的遗传相似系数为0.524~0.791, 其中卵叶山蚂蝗和圆叶舞草间的相似系数最大, 度尼山蚂蝗与异果山绿豆间的相似系数最小。在8种山蚂蝗中, 度尼山蚂蝗与其它种的相似系数均值最低, 为0.619, 说明度尼山蚂蝗与其它种的亲缘关系较远, 与聚类分析结果吻合。遗传距离分析结果与相似系数分析结果相反, 卵叶山蚂蝗和圆叶舞草间的遗传距离最小为0.209, 度尼山蚂蝗与异果山绿豆间的遗传距离最大, 为0.476。
| 表5 不同山蚂蝗种间的遗传距离(斜对线上方)和相似系数(斜对线下方) Table 5 Genetic distance (below diagonal) and genetic similarity (above diagonal) matrixes between different Desmodium species |
分子标记是作物遗传学研究和分子育种的重要工具, 相对于其它基于PCR的分子标记, SSR标记因具有数量丰富、多态性高、共显性、操作简单、重复性好等优点而被广泛应用于水稻(Oryza sativa)、拟南芥(Arabidopsis thaliana)等模式植物和玉米(Zea may)、油菜(Brassica napus)、小麦(Triticum aestivum)等大田作物的遗传学研究和分子育种[12, 25]。但对于遗传学基础薄弱的植物来说, SSR标记的开发成本较高。然而研究[26, 27]表明SSR标记在近缘物种乃至远缘物种基因组间的具有一定的通用性, 因此, 对遗传学基础较弱的物种, 物种间转移法已成为一种廉价、快捷获得SSR标记的有效途径。对1 205对绿豆(Vigna radiata)SSR标记进行筛选, 获得了603(50.0%)、883(73.3%)和983(81.6%)对分别在豇豆(V. unguiculata)、小豆(V. angularis)和饭豆(V. umbellate)中有效的SSR标记[17]。水稻和甘蔗(Saccharum officinarum)SSR标记在竹子(Bambusa spp.)中的通用性比率分别为44.9%和75%[28]; 对382对玉米SSR标记和100对甘蔗EST-SSR标记在禾本科芒属上的通用性研究[29]表明, 其可转移率分别为10.29%和13.00%。山蚂蝗是热带亚热带地区重要的豆科牧草, 当前山蚂蝗SSR标记的缺失严重限制了其遗传学研究及分子育种进展。本研究分析了92对柱花草EST-SSR标记在山蚂蝗属中的通用性, 发现柱花草EST-SSR标记在山蚂蝗中的可转移率为77.78%, 明显高于苜蓿EST-SSR标记在山蚂蝗属中的可转移率(54.17%), 说明柱花草与山蚂蝗的亲缘关系更近, 柱花草EST-SSR标记在山蚂蝗中具有较好的通用性[9]。本研究使用16对多态性EST-SSR标记对山蚂蝗遗传多样性进行分析, 发现16对EST-SSR标记可以对16份山蚂蝗种质进行有效区分, 说明本研究利用物种间转移法获得的在山蚂蝗有效的柱花草EST-SSR标记可作为山蚂蝗遗传学研究的有效工具, 为山蚂蝗的遗传多样性分析、基因发掘和鉴定、分子标记辅助育种等研究工作提供基础。
植物种质资源是新品种选育、提高农业生产力的基础资源[30], 而遗传多样性和亲缘关系的系统研究不仅与资源的收集、保存和更新密切相关, 也是种质资源创新和利用的基础[31]。传统的山蚂蝗遗传多样性研究主要以植物形态特征为依据, 如范小勇等[6]利用10个形态指标对23份山蚂蝗种质资源进行聚类分析, 最终将其分为4类, 而刘苗苗等[7]通过对山蚂蝗的16个植物学性状进行统计将37份山蚂蝗种质资源分为6类。由于山蚂蝗属植物种类繁多、植物学特征复杂多变, 有些种和亚种间的形态特征非常相近, 基于传统的形态学特征分类方法很难对这些种进行区分鉴定, 因此花粉的显微结构分析和分子标记技术被用于山蚂蝗遗传多样性和亲缘关系分析。对重庆地区6种山蚂蝗的花粉显微结构的观察比较发现, 圆菱叶山蚂蝗、宽卵叶山蚂蝗和尖叶山蚂蝗的花粉表面纹饰相似, 应归为同一亚属[32]。利用AFLP标记对来自7个种的46份山蚂蝗种质进行遗传多样性及亲缘关系分析, 最终将46份山蚂蝗种质分为6类[33]。利用ACGM和EST-SSR 两种分子标记对云贵高原的9个种46份野生山蚂蝗种质遗传多样性研究表明, 野生山蚂蝗种质间存在较高的遗传多样性[9]。但这些关于山蚂蝗遗传多样性的研究选取的材料差较大, 所以各研究结果存在一定的差异, 但都与《中国植物志》[34]中传统分类不完全一致。
本研究利用柱花草EST-SSR对来自8个种的16份山蚂蝗材料进行了遗传多样性与亲缘关系分析, 发现16份山蚂蝗种质间的相似系数为0.523~0.952, 平均0.711, 说明山蚂蝗种质的遗传多样性较为丰富。根据聚类分析在相似系数为0.730时将16份分为5类, 结果与形态学分类基本一致, 但也有类外, 如A类中包含圆叶舞草、卵叶山蚂蝗, 该结果与刘苗苗等[7]、贺欣等[9]和王春梅[32]较为相近, 但与《中国植物志》[34]分类不一致。所以, 要确定山蚂蝗种质之间的亲缘关系, 不仅要使用合适的分子标记, 还要结合各种质表型的形态多样性和生理生化多样性研究进行综合分析。
综上所述, 柱花草EST-SSR标记在山蚂蝗上具有较高的可转移性, 在8种山蚂蝗种的可转移率为63.04%~73.91%; 利用柱花草EST-SSR标记能够有效地对8种山蚂蝗进行遗传多样性及亲缘关系分析; 本研究所得通用性SSR标记将成为山蚂蝗种质资源评价及亲缘关系研究的有力工具。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|