



{kind=link}
{kind=link}
{kind=link}
{kind=link}
中国北方草原生物量年际动态
[苟晓伟 , 常生华, 程云湘, 侯扶江]
, 常生华, 程云湘, 侯扶江]
, 常生华, 程云湘, 侯扶江]
|
|


第一作者:苟晓伟(1989-),男,内蒙古镶黄旗人,在读硕士生,研究方向为草地生态学。E-mail:[email protected]
不同类型或同一类型不同区域的草地,其地上生物量在时间和空间上均存在差异,本文从不同的草地类型和时空尺度揭示植被生物量与环境因子间动态关系。结果显示,1)中国北方草原近30余年地上生物量总体呈下降趋势,水分条件较好的区域下降幅度较大,局部变化不显著,而近5年则呈持续增长趋势;2)时间序列上,年降水和年均温的交互作用对各区域草地地上生物量影响不显著;3)空间尺度上,温性草原年均温对地上生物量的影响较显著,高寒草原地上生物量的主要限制因子是降水。
Aboveground biomass in different rangeland types or same type in different regions varied with time and space which reveals the dynamic relationship between vegetation biomass and environmental factors from different rangeland types and spatial and temporal scales. The results showed that aboveground biomass in Chinese northern steppe rangeland general declined in last 30 years although consistent increased in the recent 5 years. The decline was great in areas with better water conditions and was not significant in local scale. The interaction of annual temperature and precipitation did not have significant effects on aboveground biomass in time scale. The average annual temperature of temperate rangeland had significant effects on aboveground biomass in spatial scale. The precipitation is the main limiting factor of aboveground biomass in alpine rangeland.
气候改变草原生态系统的物质环境、结构、功能[1], 中国北方草原生态系统受温度和降水的变化影响存在很多不确定性。近100年来, 全球升温幅度平均为(0.6± 0.2) ℃, 中国年平均地表气温比同期略高, 升温幅度为0.5~0.8 ℃[2]。对于中国自然生态系统来说, 气候变暖将带来显著和深远的影响, 使得人类生存环境面临巨大挑战。我国草地是陆地生态系统的主体, 约占总国土总面积的41%[3]。准确了解中国草原生态系统的变化趋势, 及时反馈全球气候变化和草地生态系统之间的关系, 将有助于监测草地资源的健康状况[4, 5, 6, 7]。
植物生物量反映能流、物流和第一性生产, 对生态系统结构和功能的形成起重要作用。因此, 国内外学者都重视对植物生物量的研究[1, 4, 6]。正确地估算我国草地植被生物量的变化, 对于研究陆地生态系统的碳循环和指导我国草地畜牧业生产具有重要意义。截至目前, 中国学者已经基于全球生物量数据库、草地资源清查资料、野外调查资料以及草地资源数据与遥感数据相结合等方法, 评价了中国草地的生物量碳库。但在大尺度生物量方面的研究, 多基于遥感信息, 较少采用实测数据, 所以存在较大不准确性[5]。
植被生物量动态与环境因子的关系因研究视角和方法不同而结论各异, 研究范围包含不同草地类型、不同时空尺度、特定的环境梯度等。草地生物量受环境因子波动的直接影响, 并且多表现为多种环境因子的综合作用, 其中关键是水热因子组合[8, 9, 10, 11]。
由于不同地域气候差异较大, 多数国内学者关注于特定地域的气候变化特征和植被生物量关系研究[12, 13]; 同时由于水热组合和土壤环境等对陆地植被影响较大[14], 且环境因子的变化必然引起群落结构和功能的变化, 进而影响群落的动态和稳定性[15]。因此, 准确揭示草地生物量波动与气候因素的关系对草地资源管理和合理利用、预测未来气候变化对草地生态系统的影响具有重要意义。然而, 目前对于草地生物量的估算及其与环境因素关系的认识仍存在争议[8]。多数研究人员采用控制试验[16]、野外调查[17]、模型模拟[18]和遥感监测[19]等方法, 研究和探讨气候变化对中国各个区域草地生态系统的影响, 但其结论缺乏整体性和系统性。而在不同尺度基础上, 探讨生物量变化与气候因子间关系的相关研究较少。
本文通过查阅近30年中国北方不同区域草地生物量的实测报道数据, 分析草地生物量和气候因子间关系, 试图揭示:1)中国北方草原地上生物量近30年动态变化与空间分布格局; 2)比较不同研究区域草地地上生物量对降水和温度的响应。
本研究所用近30年内蒙古高原、新疆和青藏高原不同类型草地地上生物量数据, 是在查阅公开发表的学术论文或出版专著基础上筛选而来, 样地均选择在地势平坦且围封的天然草地, 取样时间段均选择在植被生长旺季。年降水和年均温数据来自各地区气象站。本研究采用草地综合顺序分类法[20, 21, 22, 23], 将所涉及草地划分成暖温极干草地、微温微润草地、微温湿润草地、寒温潮湿草地、微温极干草地和寒温湿润山地草地6种类型。其中暖温极干草地、微温微润草地、微温湿润草地分布在内蒙古高原, 寒温湿润山地草地分布在天山, 寒温潮湿草地、微温极干草地则分布在青藏高原。
地上生物量与年均温、年降水的关系采用多元回归分析, 显著性检验水平为0.05。采用Kolmogorov-Smirnov检验, 分析数据是否为正态分布, 对不符合正态分布的数据使用中值计算。统计分析均采用SPSS 18.0软件进行分析。
中国北方草原不同区域多年平均地上生物量差异很大, 其变化范围为2.72~711 g· m-2(图1)。不同草地类型间地上生物量亦存在明显差异, 阿拉善暖温极干草地地上生物量只有8.741 g· m-2, 而在贵南寒温潮湿草地地上生物量高达425 g· m-2(表1)。
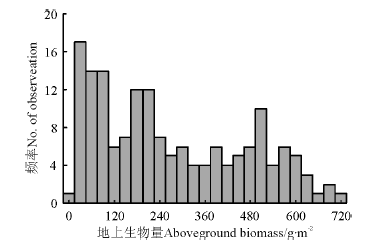 | 图1 中国北方草原地上生物量频数分布Fig.1 Frequency distribution of aboveground biomass of grassland in northern China |
| 表1 不同区域地上生物量均值、最大值和最小值 Table 1 Aboveground biomass of different regions in northern China from 1980 to 2000 |
阿拉善定点监测数据显示, 1993-2008年, 荒漠草地地上生物量呈逐年下降趋势, 年均降幅达6.11 kg· hm-2(图2a); 综合锡林郭勒盟微温微润草地的监测数据, 1984-2013年, 地上生物量平均以每年42.62 kg· hm-2的速度下降; 锡林浩特市草地地上生物量监测数据表明, 地上生物量以每年24.33 kg· hm-2的幅度下降(图2b); 但是西乌珠穆沁旗的产草量逐年略有增加(图2c)。1984-1995年, 呼伦贝尔的微温湿润草地(鄂温克旗)年地上生物量平均以111.7 kg· hm-2的速度递增(图2d)。天山的寒温湿润山地草地, 2000-2010年, 地上生物量年均增速为500.1 kg· hm-2(图2g)。在青藏高原, 2006-2011年, 若尔盖寒温潮湿草地地上生物量以每年357.1 kg· hm-2的速度下降(图2h); 1984-2009年, 贵南草地地上生物量呈较小波动趋势(图2i); 1994-2004年, 格尔木微温极干草地地上生物量以每年22.81 kg· hm-2的速度上升, 但趋势不明显(图2 j)。
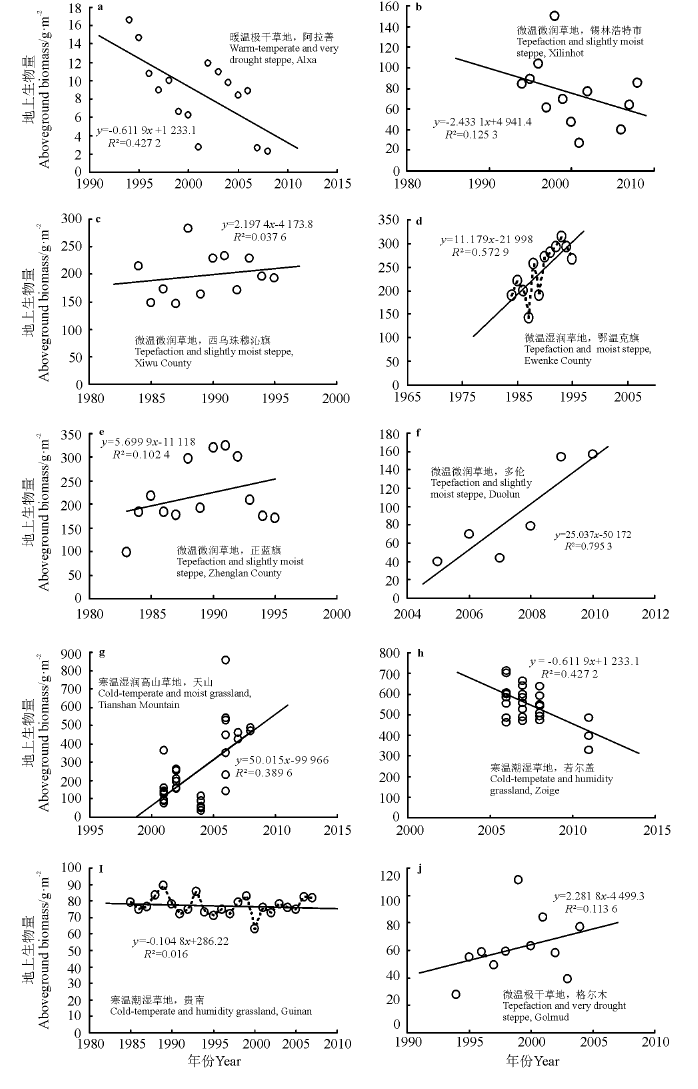 | 图2 近30年间不同草地类型地上生物量的变化Fig.2 Spatial distribution of grassland biomass in northern China |
不同区域的响应存在差异, 其中锡林浩特微温微润草地(R2=0.71, P< 0.05)和天山寒温湿润山地草地(R2=0.19, P< 0.05)年降水对地上生物量影响显著, 而在贵南寒温潮湿草地(R2=0.89, P< 0.01)地上生物量与年均温的作用显著相关, 其它区域草地的地上生物量与气候因子都不相关。
温性草原地上生物量与年降水的正相关关系不显著(R2=0.244 1, P> 0.05)(图3a), 但是与年均温显著负相关(R2=0.776 7, P< 0.01)(图3b); 高寒草原的结论正好相反, 其地上生物量与年降水量有显著的正相关关系(R2=0.930 3, P< 0.01)(图3c), 但与年均温的负相关关系不显著(R2=0.378 6, P> 0.05)(图3d)。整个北方草原范围内, 草地地上生物量与年降水有显著的正相关关系(R2=0.356 9, P< 0.05)(图4)。
 | 图3 温性草原(暖温极干草地、微温微润草地、微温湿润草地)和高寒草原(寒温潮湿草地、微温极干草地)地上生物量与气候因子的回归关系Fig. 3 Relationships of biomass with climatic factors for temperate steppe and alpine grasslands in northern China |
 | 图4 中国北方草原地上生物量与年降水量的关系Fig. 4 The relationship between annual precipitation and grassland aboveground biomass in northern China |
本研究中国北方草原1980-2010年平均地上生物量波动范围是2.72~711 g· m-2, 而Yang等[31]和Piao等[32]报道中国北方草地生物量分别为104.8和97~102 g· m-2(1982-1999年); 原因可能是本研究所用数据在时间序列范围更大, 。全球草原平均地上生物量378.4 g· m-2[31], 中国北方草原与之相比是一个低水平产草量的区域。
从时间序列上看, 中国北方不同类型草原生物量变化速度不同, 其中环境条件较好区域的地上生物量变化速度较大, 锡林郭勒盟微温微润草地在1984-2013年呈下降趋(42.62 kg· hm-2· a-1)、鄂温克旗微温湿润草地在1984-1995年呈上升趋(111.7 kg· hm-2· a-1)以及若尔盖寒温潮湿草地在2006-2011年呈下降趋势(357.1 kg· hm-2· a-1), 而环境较差区域的地上生物量年际间变化速度明显小于环境条件好的地区。原因可能是草地植被在不同生态环境下对生物量分配的差异造成的, 水分和光照是植物生长的限制因素, 植被通常在适宜的环境下会分配更多的地上生物量, 所以环境条件较好的微温湿润草地、微温微润草地和寒温潮湿草地的地上生物量受降水和温度的影响更为明显, 变化范围更大。
总体上, 从1980年开始我国草原产草量总体下降, 但生物量没有明显的变化趋势, 该结果与内蒙古温带草地[33]和青藏高原高寒草地的长期定位观测结果[34]一致, 即在过去的30多年尽管温度显著升高, 但中国北方草地的地上生物量并没有显著变化, 中国北方草地生产力保持在一个稳定的状态。
降水在空间尺度上被认为是影响草地生态系统生物量变化的主要因素[35, 36, 37, 38], 牧草的生长和产量受降水和温度共同影响[39], 在中国陆地生态系统中, 生产力与降水的年际波动间存在密切关系, 群落初级生产力随着年降水的增多也相应增加[40]。但是, 在时间尺度上二者是否存在显著的相关性, 目前仍因研究结论不同而存有争议[41]。本研究中, 只有锡林浩特、天山和贵南3个区域的地上生物量与年降水量和年均温的相关性显著, 其它研究认为草地地上生物量与气候因子没有显著的相关性[42]。这是由于局部地区降水和温度对生物量的影响复杂, 草地地上生物量变化受多种因子影响。比如, 有研究表明放牧对中国北方草原地上和地下生物量的形成有很大的抑制作用, 放牧约降低总生物量的60%[43], 所以在某些特定区域内降水和温度对生物量的影响会被弱化。整体上气候因子对生物量影响是显著的, 但小范围内却没有什么规律。目前, 气候因子对草地植被群落的生产力的影响仍因研究结论的不同而存有争议, 更多的研究结论还有待从不同角度进行深入研究[38, 40, 44]。
大气-植被互作模式对内蒙古半干旱草原净初级生产力和生物量的模拟表明, 地上生物量随着降水的增加而上升, 随着温度升高而下降[45]。本研究在时间序列的基础上扩展到空间尺度, 发现年降水量对整个中国北方草地地上生物量有显著影响(图4)。在温性草原区域内, 草地生物量的主要影响因素是温度, 随着温度的升高草地地上生物量呈下降趋势。之前很多研究都表明, 草地地上生物量随着温度增加和水分的降低而减少[46, 47, 48]。干旱的条件下植物光合作用可能被高温约束, 所以在内蒙古微温微润草地高温导致光合速率的下降进而影响生物量。更高的温度会导致蒸发增加, 加剧干旱和减少生物量[49]。高寒草原草地生物量的主要影响因子是降水, 草地生产力随着降水的增加呈明显的增加趋势。基于三江源区兴海县46年的气候资料分析[50], 降水量是影响草地生产潜力的限制因子。
基于国内外学者的研究表明, 受气候等因素的影响我国北方草原地上生物量近30年各地区变化不一, 总体上说保持在一个稳定状态。尺度研究对于草地生物量的研究很有意义, 在时间序列上气候因子对草地生物量的影响被弱化, 而在空间尺度上由于气候条件差异较大, 降水量和温度对草地生物量影响显著。中国北方草原中温性草原生物量的主要限制因子是温度, 相反限制高寒草原生物量的主要气候因子是降水量。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|