





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
抗裂荚箭筈豌豆荚果发育动态及其腹缝线结构
引用本文
董德珂, 韩云华, 李东华, 王彦荣, 刘志鹏. 抗裂荚箭筈豌豆荚果发育动态及其腹缝线结构. 草业科学, 2017,34(11):2289-2294
Dong De-ke, Han Yun-hua, Li Dong-hua, Wang Yan-rong, Liu Zhi-peng. Study on pod development and ventral suture structure of shattering-resistant common vetch. Pratacultural Science,2017,34(11): 2289-2294
Permissions
Dong De-ke, Han Yun-hua, Li Dong-hua, Wang Yan-rong, Liu Zhi-peng. Study on pod development and ventral suture structure of shattering-resistant common vetch. Pratacultural Science,2017,34(11): 2289-2294
Copyright©2017, 《草业科学》编辑部
抗裂荚箭筈豌豆荚果发育动态及其腹缝线结构
第一作者:董德珂(1991-),男,山东平邑人,在读硕士生,主要从事草类作物遗传育种研究。E-mail:[email protected]
摘要
易裂荚箭筈豌豆( Vicia sativa)的荚果开裂主要是由离层和细胞失水产生的机械拉力导致的,但是在抗裂荚的箭筈豌豆中的组织结构尚不明确。该研究以抗裂荚的135号箭筈豌豆( V. sativa subsp. sativa No.135)为对象,对其荚果在发育过程中的形态特征、水分含量、发芽率及腹缝线横截面解剖结构等的动态变化进行观察分析,以探讨抗裂荚箭筈豌豆荚果的抗裂机理,以及为生产中确定其种子收获的适宜时间提供理论依据。结果显示,1)135号箭筈豌豆约在盛花后26 d荚果变为棕黑色,荚果的大小、干重已经达到最大值,含水量较低,发芽率较高且硬实率较低。2)135号箭筈豌豆腹缝线处夹在两个维管束之间的薄壁细胞没有分化成离层,而是分化成与维管束细胞一样的厚壁细胞,将两个维管束连接起来,形成由厚壁细胞组成的整体。综上所述,盛花后26 d荚果变为棕黑色时是135号箭筈豌豆的适宜收获时间,离层的丢失和分化形成的厚壁细胞是其荚果抗裂的主要原因。
关键词:
箭筈豌豆; 裂荚; 离层; 适宜收获时间; 腹缝线
中图分类号:S551+.9
文献标志码:A
文章编号:1001-0629(2017)11-2289-06
doi: 10.11829/j.issn.1001-0629.2016-0592
Study on pod development and ventral suture structure of shattering-resistant common vetch
Abstract
The abscission layer cells and machine tensile generated by cellular water loss were the main causes of pod shattering in shattering-susceptible common vetch ( Vicia sativa), but the organizational structure is unclear. This study, using shattering-resistant V. sativa subsp. sativa No.135 as the research material, analyzed its dynamic changes in pod morphology, moisture content, germination rate and anatomical structure of ventral suture cross section during pod development to explore the pod shattering-resistant mechanism and provide a theoretical basis for determining optimal harvest time. The results elucidated two main points: 1) When the pods of V. sativa subsp. sativa No.135 turned to brown-black at about 26 days after peak anthesis, they reached their maximum size and dry weight; when the germination rate was high, the hard seed percentage and the water content were low. 2) The parenchyma cells between the two vascular bundles were not differentiated into abscission layers, but differentiated into sclerenchymatous cells and connected the vascular bundles, wholly formed by the parenchyma cells. The results showed that the optimum harvest time was 26 days after peak anthesis. The key cause of the shattering-resistant trait lies in the loss of layer cells and the parenchyma cells differentiated into the sclerenchymatous cells.
Keyword:
V. sativa; pod shattering; layer cells; optimum harvest time; ventral suture
箭筈豌豆(Vicia sativa)是豆科(Leguminosae)野豌豆属(Vicia)一年生的闭花授粉植物, 具有适应性广、抗寒性强、营养价值高等优良特性, 是重要的牧草和绿肥作物, 缓解了我国高海拔地区冬季饲草匮乏的问题, 在我国草地农业系统中发挥着不可替代的作用[1, 2, 3, 4, 5, 6]。但是, 箭筈豌豆在荚果成熟时的开裂现象会导致种子的严重损失, 不利于箭筈豌豆的大面积推广。兰州大学草地农业科技学院从49个国家收集了500余份箭筈豌豆种质, 经过多年的筛选和驯化, 选育出了抗裂荚的135号箭筈豌豆, 具有裂荚率低、抗寒性强、地上生物量和种子产量高等优点, 有望培育成高海拔地区反刍动物的优质牧草新品系。
种子适宜收获时间的确定对牧草种子生产具有重要的意义, 收种时间不当往往是造成种子品质差、产量低的主要原因[7]。为了避免因收获时间不当带来的种子产量的损失, 从19世纪60年代起, 国内外的一些学者开始致力于研究种子的适宜收获时间[8]。对于不同物种, 确定其适宜收获时间的方法各有差异, 总体来说, 种子适宜收获确定的主要依据有荚果及种子的发育时间[9, 10]、外部形态特征[10, 11]、干鲜重和含水量等生理指标[12, 13]以及发芽率[9, 13]等。
荚果本身的组织结构对裂荚起着决定性的作用[14], 研究表明, 豆科植物荚果开裂的起始部位是腹缝线[15]。目前, 关于裂荚的研究在大豆(Glycine max)和拟南芥(Arabidopsis thaliana)中相对成熟, 主要集中在荚果裂荚区的解剖结构, 其中荚果开裂及抗裂荚的重要结构包括离层、内果皮的内厚壁组织以及维管束等[16, 17, 18]。在大豆裂荚的最新研究中, 对比了野生型和栽培型大豆的腹缝线结构差异, 指出经过5 000多年的驯化, 栽培大豆中位于离层上方的纤维帽细胞的数量和细胞壁的厚度远高于野生型大豆, 大大增强了其抵抗由离层细胞释放的水解酶的能力, 表明纤维帽细胞是大豆抵抗裂荚的关键结构[18]。对中度裂荚的箭筈豌豆兰箭3号荚果的解剖结构发现, 箭筈豌豆中没有纤维帽细胞, 取而代之的是外部果瓣缘细胞外侧异常加厚的细胞壁, 推测其是箭筈豌豆荚果抗裂的关键结构, 同时离层和细胞失水产生的机械拉力是其裂荚的主要原因[19], 但是抗裂荚箭筈豌豆的组织结构尚不明确。因此, 本研究以抗裂荚的135号箭筈豌豆为材料, 对其各发育时期的荚果腹缝线进行解剖结构观察, 并与中度裂荚的兰箭3号进行对比分析, 试图找到其他与裂荚抗性相关的结构, 为进一步研究箭筈豌豆的裂荚机理奠定基础。
1 材料和方法
1.1 试验材料
本研究所用材料兰箭3号和135号箭筈豌豆均由兰州大学草地农业科技学院、草地农业生态系统国家重点实验室提供。
1.2 试验设计
试验地点设在甘肃省兰州市兰州大学榆中校区的试验地(35.57° N, 104.09° E)。试验中选取各份材料的正常饱满的种子, 种植在面积为1 m× 25 m的试验小区, 采用单行单株种植的方法, 株距为50 cm, 每份种质3次重复, 灌水等其他条件保持相同。在盛花期时标记135号箭筈豌豆完全开放的花朵(以旗瓣完全展开为完全开放)。
1.3 取样与处理
盛花期标记的花朵, 分别在盛花期后5、10、15、20、23、26、29、32 d进行取样。各取样时期在各小区中随机取10个正常的荚果, 用来测定荚果及种子的形态学、生理学等指标。从每个小区另取若干个荚果放入FAA固定液(70%乙醇∶ 冰乙酸∶ 甲醛=9∶ 0.5∶ 0.5)中固定, 并保存在4 ℃冰箱中, 用于制备冷冻切片。荚果成熟时, 取若干个兰箭3号和135号的荚果, 用于测定其裂荚力和裂荚率。
1.4 荚果裂荚力、裂荚率的测定
荚果裂荚力、裂荚率参照董瑞等[20]的方法测定。
1.5 荚果形态学特征
把荚果分别放在平板上, 旁边放置1 cm的比例尺, 并拍照, 再用Digimizer 4.2软件测量荚果的长和宽; 利用游标卡尺测量荚果的厚度, 精确到0.01 mm。
1.6 种子和荚果的生理学特征
把每个荚果的荚皮和种子进行分离, 并用电子天平称量其鲜重, 精确到0.001 g。然后将其放在80 ℃的烘箱中烘干至恒重并称量, 精确到0.001 g, 计算含水量。
含水量=(鲜重-干重)/鲜重× 100%。
1.7 种子发芽情况
根据GB/T 2930.4-2001《牧草种子检验规程发芽试验》[21]进行箭筈豌豆种子的标准发芽试验。发芽床采用滤纸法, 每个取样期设3次重复, 每个重复用50粒种子。在20 ℃恒温培养箱(12 h光照, 12 h黑暗)内培养, 每24 h补充适当水分。以胚根突出种皮视为萌发, 第5天初次统计, 末次统计为第14天, 14 d后不吸胀的种子统计为硬实。
1.8 腹缝线横截面解剖结构观察
取出FAA固定液中的荚果, 截取腹缝线中间约3 mm的一段, 然后通过70%(1 h)、80%(1 h)、95%(1 h)、95%(1 h)、100%(1 h)、100%(1 h)、100%(1 h)的乙醇进行梯度脱水。利用Leica CM3050冷冻切片机切片, 厚度为5 μ m, 染液为甲苯胺蓝, 中性树胶封片, OLYMPUS BX51显微镜拍照。
1.9 数据分析
利用SPSS 17.0对兰箭3号和135号箭筈豌豆荚果裂荚力和裂荚率的差异进行t检验(P< 0.05), 利用Excel 2016计算出各指标的平均值和标准误, 并作图。
2 结果与分析
2.1 兰箭3号和135号箭筈豌豆荚果裂荚力、裂荚率的对比
兰箭3号的裂荚率为50%左右, 显著高于135号种质(裂荚率为2.5%左右)(图1)。135号在水平放置和垂直放置时开裂所需的机械力(即裂荚力)显著高于兰箭3号。
 | 图1 兰箭3号和135号箭筈豌豆荚果裂荚率和裂荚力 注:* 表示同一指标两个种质间差异显著(P< 0.05)。Fig. 1 Pod-shattering rate and mechanical force of Lanjian No.3 and No.135 Note:* indicate significant difference of the same parameter between two germplasms at the 0.05 level. |
2.2 135号箭筈豌豆荚果的形态学特征
135号箭筈豌豆荚果的颜色随生长时间的推移而发生显著的变化(图2)。盛花后5-20 d荚果颜色为绿色; 盛花后23 d荚果颜色变为棕黄色; 随着荚果的进一步发育, 盛花后26-32 d荚果颜色变为棕黑色。
 | 图2 135号箭筈豌豆荚果发育过程中荚果的颜色Fig. 2 Pod color during pod development of V. sativa subsp. sativa No.135 |
135号箭筈豌豆荚果的长度和宽度在盛花后0-15 d时迅速增加, 并于盛花后15 d左右达到最大, 随后基本保持不变(图3)。荚果的厚度在盛花后0-23 d时持续增长, 于盛花后23 d左右达到最大。
 | 图3 135号箭筈豌豆荚果发育过程中荚果的长度、宽度和厚度Fig. 3 Pod length, width and thickness during pod development of V. sativa subsp. sativa No.135 |
2.3 135号箭筈豌豆荚皮和种子的生理学特征
135号箭筈豌豆荚皮鲜重在盛花后0-20 d呈逐渐增加的趋势, 大约在盛花后20 d达到最大值, 随后逐渐减小, 至盛花后29-32 d达到最小值; 荚皮含水量在盛花后5-29 d呈逐渐减小的趋势, 至盛花后29-32 d达到最小值, 盛花后26 d约为30%; 荚皮的干重在盛花后0-23 d逐渐增加, 在盛花后20 d左右达到最大值, 随后保持不变(图4)。
 | 图4 135号箭筈豌豆荚果发育过程中荚皮鲜重、干重和含水量Fig. 4 Pod wall fresh weight, dry weight and moisture content during pod development of V. sativa subsp. sativa No.135 |
135号箭筈豌豆种子鲜重在盛花后0-23 d呈逐渐增加的趋势, 随后逐渐减小, 在盛花后29-32 d趋于稳定; 种子含水量在盛花后10-29 d呈逐渐减小的趋势, 盛花后29-32 d达到最小值, 在此期间无明显变化, 盛花后26 d约为30%; 种子干重在盛花后0-10 d时开始增长, 但是增长缓慢, 盛花后10-26 d是种子干重的迅速增长期, 并在盛花后26 d左右达到最大值, 随后保持不变(图5)。
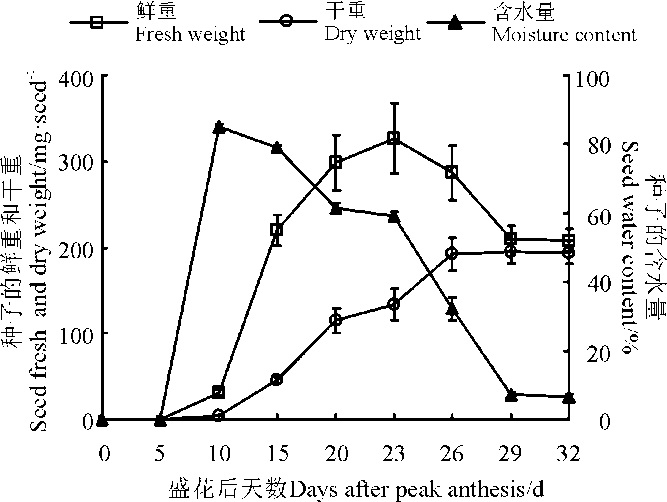 | 图5 135号箭筈豌豆荚果发育过程中种子鲜重、干重和含水量Fig. 5 Seed fresh weight, dry weight and moisture content during pod development of V. sativa subsp. sativa No.135 |
2.4 135号箭筈豌豆不同发育时期种子的发芽情况
盛花后15 d的135号箭筈豌豆种子发芽率为0, 盛花后20 d发芽率迅速提高, 至盛花后23 d发芽率达到最大值, 随后发芽率逐渐降低(图6)。种子的硬实率随着发育时间逐渐增高, 在盛花后32 d左右达到最大值, 约为40%。
 | 图6 135号箭筈豌豆发育过程中种子发芽率和硬实率Fig. 6 Seed germination rate and hand seed rate during development of V. sativa subsp. sativa No.135 |
2.5 135号箭筈豌豆腹缝线横截面的冷冻切片
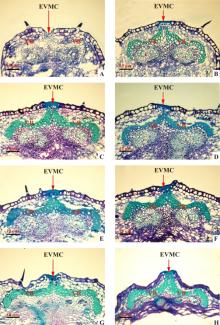
盛花后5 d, 135号箭筈豌豆外部果瓣缘细胞的外侧细胞壁开始加厚, 但不明显; 维管束细胞的细胞壁还没有加厚的迹象, 但是已经形成了维管束的雏形(图7A)。盛花后15 d, 外部果瓣缘细胞的外侧细胞壁明显加厚, 并融合为一个整体; 维管束细胞的细胞壁也明显加厚, 夹在两个维管束中间的是一些薄壁细胞(图7B), 前期的研究结果显示, 这些薄壁细胞在兰箭3号箭筈豌豆中会分化成离层细胞。盛花后15 d, 外部果瓣缘细胞的外侧细胞壁和维管束细胞的细胞壁继续加厚, 并且维管束中间的薄壁细胞有一部分分化成跟维管束细胞一样的厚壁细胞, 但是中间仍有部分细胞隔开了两部分维管束(图7C)。盛花后20 d, 外部果瓣缘细胞的外侧细胞壁和维管束细胞的细胞壁的厚度不再发生明显变化, 由维管束中间的薄壁细胞分化而来的厚壁细胞将维管束两部分连接在一起(图7D)。盛花后23-26 d, 内、中、外果皮的薄壁细胞逐渐失水皱缩, 但是荚果的两个果瓣并没有开裂的迹象, 维管束和外部果瓣缘细胞由于存在加厚的细胞壁, 形态上没有发生变化(图7E、7F、7G、7H)。
 | 图7 135号箭筈豌豆荚果发育过程中荚果腹缝线横截面的冷冻切片 注:A~H表示盛花后5、10、15、20、23、26、29、32 d; EVMC, 外部果瓣缘细胞; VB, 维管束。Fig.7 Frozen slices of pod ventral suture cross-sectional during pod development of V. sativa subsp. sativa No.135 Note:A~H stand for 5, 10, 15, 20, 23, 26, 29, 32 days after peak anthesis. EVMC, external valve margin cells; VB, vascular bundle. |
3 讨论
在实际生产过程中, 确定种子适宜收获时间至关重要, 适宜收获时间的判定方法有很多, 其中荚果和种子外部形态变化及成熟状态的指标被认为是最有效且便捷的方式, 这种方法既可以避免测定种子生理指标和种子品质花费的大量时间, 又不用破坏种子的内部
结构, 因此, 形态学指标在实际生产中应用的意义较大[7, 8]。毛培胜等[9]在河北省承德市利用荚果和种子的生长发育时间来确定老芒麦(Elymus sibiricus)种子的收获时间, 认为在盛花后26~27 d收获可以获得该地区较高的种子产量。Elias和Copeland[10]认为荚果和种子的颜色等外部形态特征以及种子含水量是判断加拿大油菜(Brassica napus)种子收获时间确定的精确简捷的指标。Fraser等[12]研究表明, 种子含水量是大豆生理收获期的准确指标。本研究发现, 盛花后26 d时其荚果变为棕黑色, 经过测定此时荚果的长度、宽度、厚度, 荚皮和种子的干重已经达到最大值, 含水量较低, 发芽率相对较高且硬实率较小, 说明此时已经完成了生理成熟且硬实还没有完全形成, 因此, 推断盛花后26 d荚果变为棕黑色时是135号箭筈豌豆种子的适宜收获时间, 本研究为其种子生产提供了理论依据。
离层的丢失和分化形成的厚壁细胞是135号箭筈豌豆荚果抗裂荚的关键因素。通过对比易裂荚种质兰箭3号[19]和抗裂荚种质135号箭筈豌豆荚果的腹缝线解剖结构发现, 二者最大的区别在于离层的有无, 其他结构均无明显差异。诸多研究表明, 离层是豆科植物荚果开裂不可或缺的关键结构[16]。前期对易裂荚种质兰箭3号荚果的解剖结构发现, 离层是由夹在两个维管束中间的薄壁细胞组成的, 离层细胞释放水解酶, 使细胞解体, 是荚果开裂的起始部位, 也是裂荚的首要原因[19]。但是本研究中, 抗裂荚的135号箭筈豌豆荚果在发育过程中并没有产生离层, 夹在两个维管束中间的薄壁细胞不是分化成离层, 而是分化成了与周围维管束一样的厚壁细胞, 并把两个维管束紧紧连接在一起, 极大地增强了组织的支撑能力, 另一方面离层在发育后期会产生水解酶使细胞裂解, 因此, 离层的丢失也避免了细胞裂解的发生。值得一提的是, 在兰箭3号荚果的解剖结构中, 外部果瓣缘细胞是其荚果抗裂的一个关键结构, 在135号箭筈豌豆种质中也存在该结构, 表明外部果瓣缘细胞是箭筈豌豆荚果抗裂不可或缺的存在。综上所述, 离层的丢失和分化形成的厚壁细胞是135号箭筈豌豆荚果抗裂的主要原因。
The authors have declared that no competing interests exist.
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|