
{kind=link}
{kind=link}
CO2浓度升高对不同秋眠类型苜蓿生长发育的影响
引用本文
翟晓朦, 张晓波, 王铁梅, 关潇. CO2浓度升高对不同秋眠类型苜蓿生长发育的影响. 草业科学, 2017,34(3):523-531
Zhai Xiao-meng, Zhang Xiao-bo, Wang Tie-mei, Guan Xiao. Effects of elevated CO2 on growth and yield of different fall dormancy alfalfa . Pratacultural Science,2017,34(3): 523-531
Permissions
Zhai Xiao-meng, Zhang Xiao-bo, Wang Tie-mei, Guan Xiao. Effects of elevated CO2 on growth and yield of different fall dormancy alfalfa . Pratacultural Science,2017,34(3): 523-531
Copyright©2017, 《草业科学》编辑部
《草业科学》编辑部
CO2浓度升高对不同秋眠类型苜蓿生长发育的影响
第一作者:翟晓朦(1990-),女,江苏兴化人,助理研究员,硕士,研究方向为草地资源的研究与利用。E-mail:[email protected]
摘要
为了分析3种秋眠类型苜蓿( Medicago sativa)(秋眠型苜蓿 Maverick、半秋眠型苜蓿ABI700和极非秋眠型苜蓿UC-1465)在不同CO2浓度下的生长发育情况,本研究运用人工气候箱和开顶式气室动态观测了3种秋眠类型苜蓿的种子发芽特性、生长发育和产量大小。主要结果:1)550、700 μmol·mol-1CO2浓度处理能显著提高3种秋眠类型苜蓿种子的发芽率、发芽势和发芽指数( P<0.05),缩短平均发芽天数( P<0.05)。其中550 μmol·mol-1浓度处理下苜蓿种子的发芽率、发芽势最高。2)高CO2浓度促进3种秋眠类型苜蓿生育期提前3~6 d,且CO2浓度越高生育期越提前。3)高CO2浓度能显著促进3种秋眠类型苜蓿株高、叶面积和单株干重增加( P<0.05)。高CO2浓度处理后UC-1465的单株鲜重比Maverick高19.33%~21.76%,比ABI700高12.74%~13.11%;单株干重比Maverick高14.85%~26.77%,比ABI700高12.31%~14.58%,高CO2浓度处理后极非秋眠型苜蓿UC-1465生长优势进一步提高。4)CO2浓度对苜蓿茎粗无显著影响( P>0.05),但促进秋眠型苜蓿Maverick、半秋眠型苜蓿ABI700茎粗增大。上述结果表明,在一定浓度范围内,苜蓿种子的发芽率、发芽指数、发芽势、株高、叶面积、生物量等对CO2的响应存在浓度效应。可以预测,未来大气CO2浓度升高的对苜蓿生长有一定的促进作用,具有一定的应用潜力。
关键词:
CO2浓度; 秋眠型苜蓿; 种子发芽; 株高; 叶面积; 茎粗; 产量
中图分类号:S541.9
文献标志码:A
文章编号:1001-0629(2017)3-0523-09
doi: 10.11829/j.issn.1001-0629.2016-0132
Effects of elevated CO2 on growth and yield of different fall dormancy alfalfa
Abstract
This study was conducted to determine the growth and development of three types of fall dormancy alfalfa (fall dormancy alfalfa Maverick, semi-fall dormancy alfalfa ABI700, and non-fall dormancy alfalfa UC-1465) under elevated CO2 concentration. We used an artificial climate box and open top chamber dynamic and observed seed germination, growth, and yield of three types of fall dormancy alfalfa. The results showed the following: 1) The germination rate, germination potential, germination index, and mean germination time of all three types fall dormancy alfalfa were promoted significantly when CO2 concentrations reached 550-700 μmol·mol-1. 2) Under high CO2 concentration, the growth period of the three types of alfalfa increased with CO2 concentration after 3~6 d. 3) The plant height, leaf area, fresh weight, and dry weight increased significantly under high CO2 concentration ( P<0.05). The fresh weight of single plant of UC-1465 was greater than Maverick and ABI700 by 19.33%~21.76% and 12.74%~13.11%, respectively, after high CO2 concentration treatment. The dry weight per plant of UC-1465 was also greater than Maverick and ABI700 by 14.85%~26.77% and 12.31%~14.58%, respectively, after high CO2 concentration treatment, while the growth advantage of the non-fall dormancy alfalfa UC-1465 was further improved. 4) CO2 concentration had no significant effect on the stem diameter except at flowering ( P>0.05). The stem diameters of Maverick and ABI700 during the whole growth period were larger than those of the control group under high CO2 concentration. These results suggest that the growth potential of the non-fall dormancy alfalfa and semi-fall dormancy alfalfa would be greater in the future under conditions of elevated atmospheric CO2 concentration.
Keyword:
CO2 concentration; fall dormancy of alfalfa; seed germination; plant height; leaf area; stem thickness; yield
政府间气候变化专门委员会(IPCC)指出, 由于一次性能源如煤、石油、天然气等矿物燃料的消费结构比重加大, 大气中“ 温室气体” 尤其是CO2气体浓度急剧升高。据2007年第4次发表正式的“ 气候变迁评估报告” 中指出, 目前全球大气CO2浓度达到了350~379 μ mol· mol-1, 21世纪中叶估计将达到550 μ mol· mol-1左右, 21世纪末估计将达到700 μ mol· mol-1左右[1]。
CO2加富试验是模拟全球气候变化的控制试验, 多数试验证明CO2浓度升高对植物的物质积累具有正效应[2, 3], 但也有其它研究结果不一致[4, 5, 6, 7]。高CO2浓度对植物的形态结构有很大的影响, 大量试验证明高CO2浓度可以促进植物光合作用, 提高地上部生物量、株高、叶面积、分枝数、茎粗等指标, 且随CO2浓度升高而升高[8, 9], 使干重大大增加[10, 11, 12]。CO2加富试验发现CO2浓度升高至760.1 μ mol· mol-1冬小麦(Triticum aestivum)的地上部、地下部干重明显增加(P< 0.05), 穗重, 穗数增加, 根系发达, 根长、根密度明显上升(P< 0.05)[11]。CO2浓度升高还促进水稻(Oryza sativa)生育期提前[13], 白花蝇子草(Silene pratensis)的花期也提前了0.5 d左右[14]。此外, CO2浓度升高是否会对植物种子发芽产生促进或抑制的作用, 目前国内外只有很少的文献涉及到此方向[15]。CO2是植物的“ 绿色碳汇” , 其浓度升高对植物的生命活动, 生长发育和产量有较大的影响, 最终影响自然生态系统和农业生态系统[16, 17]。
紫花苜蓿(Medicago sativa)是世界上栽培历史悠久、栽培面积最广的豆科牧草之一, 堪称“ 牧草之王” 。我国苜蓿品种繁多, 不同秋眠型苜蓿生态适应各不相同。秋眠性(fall dormancy)是苜蓿秋末刈割后对光温变化的不同响应的特征, 秋眠级越高, 苜蓿的茎越直立, 生长越旺盛, 产量越高, 但越冬性差[18]。本研究分析了不同CO2浓度下, 不同秋眠类型苜蓿生长发育和产量的变化, 对指导我国苜蓿产业发展具有重要的意义。
1 材料与方法
1.1 供试材料
试验采用美国标准对照秋眠级的苜蓿品种:Maverick(简称FD1)、ABI700(简称FD6)和UC-1465(简称FD11), 来自北京林业大学。苜蓿品种名称及其秋眠级见表1。
| 表1 苜蓿品种秋眠级及来源 Table 1 Fall dormancy and origins of alfalfa |
1.2 研究方法
1.2.1 种子发芽试验 试验在CO2人工气候箱中进行。设置白天12 h, 光强15 000 lx; 夜晚12 h, 完全黑暗, 恒温25 ℃, 恒湿60%[19]。CO2浓度处理分别为550、700 μ mol· mol-1以及大气CO2浓度作为对照CK。试验按《农作物种子检验规程》方法操作[20]。以种子胚根突破种皮2 mm为发芽, 持续8 d对发芽情况进行统计。每个处理4次重复, 最后取其平均值[21]。
发芽率=第8天发芽种子总数/供试种子总粒数× 100%。
发芽势=第4天发芽种子粒数/供试种子总粒数× 100%。
发芽指数(GI)=∑ (Gt/Dt),
式中:Gt为在不同天数的发芽率; Dt为发芽天数。
平均发芽天数=∑ (Gt× Dt)/∑ Gt
式中:Gt、Dt与发芽指数公式中相同[19]。
1.2.2 生长发育试验 试验地位于北京市赵全营镇中国环境科学研究院试验站, 地理位置为38° 39'27.97″ N, 104° 04'58.66″ E, 海拔28.5 m, 属暖温带半湿润气候。主要使用的设备为六边形无色透明玻璃结构的开顶式气室(Open-top chamber), 气室内供气装置为CO2 钢瓶, 纯度为99%。分别设置550、 700 μ mol· mol-1两个试验组, 大气环境本底CO2浓度350 μ mol· mol-1为对照组(CK), 每组均设4个重复, 每个重复4个花盆, 每个花盆10株苜蓿, 整个试验期内每盆苜蓿每天等量浇水一次。
1.2.3 指标测定方法
生育期:按《牧草-草地农业科学》记载标准[22], 鉴定苜蓿分枝期、初花期、盛花期、成熟期等。记载不同处理下各生育期的日期, 计算播种后各生育期的天数(d)。
叶面积:选取发育一致的健康苜蓿枝条顶部向下第2片中间展开叶, 每个处理取完全展开叶30片, 每个生育期各测一次。
株高:长出子叶后, 每重复随机测3株, 苗期7 d测一次, 分枝期、初花期、盛花期和成熟期各测一次。
茎粗:长出子叶后, 每重复随机测3株, 分枝期、初花期、盛花期和成熟期各测一次。
草产量:初花期时每个重复选8个花盆, 每盆选取5株具有代表性的植株, 剪取离土面大约1 cm的地上部, 用鼓风干燥箱于105 ℃下杀青5 min, 然后在80 ℃烘干至恒重, 称量干重。
1.3 数据分析
试验数据采用Excel 2010软件进行初步处理, 试验结果以平均值± 标准差表示。利用SPSS 18.0统计软件的ANOVA程序进行方差分析, 差异显著时采用LSD方法进行多重比较。
2 结果与分析
2.1 CO2浓度对苜蓿种子发芽的影响
CO2浓度升高显著促进苜蓿种子发芽率(P< 0.05)(表2), 其中未发芽的种子处于吸胀状态, 550 μ mol· mol-1处理下苜蓿的发芽率最高, 每种苜蓿在不同处理下种子发芽率大小排序均为550 μ mol· mol-1> 700 μ mol· mol-1> CK。
CO2浓度升高显著促进发芽势(P< 0.05)(表2), 700和550 μ mol· mol-1处理下, FD1发芽势分别较CK提高了15.82%和25.31%; FD6分别提高了15.88%和34.11%; FD11分别提高了7.51%和21.95%。CO2浓度达到550 μ mol· mol-1时, 3种秋眠类型苜蓿的种子发芽势显著高于对照组(P< 0.05)。
| 表2 不同CO2浓度对种子发芽率、发芽势、发芽指数和平均发芽天数的影响 Table 2 Seed germination rate, germination potential, germination index, and the mean germination days of alfalfa under different CO2 concentrations |
CO2浓度对发芽指数的影响显著(P< 0.05)。700、550 μ mol· mol-1处理下, FD1发芽指数分别提高了43.50%、42.31%; FD6分别提高了25.87%、54.07%; FD11分别提高了17.20%、40.80%。发芽指数能灵敏地表现种子活力, 550 μ mol· mol-1组FD6和FD11发芽指数达到最大, 种子活力指数也最高, 与FD1发芽指数差异均显著(P< 0.05)。
此外, CO2浓度升高显著缩短了苜蓿的发芽时间(P< 0.05), 700 μ mol· mol-1处理的苜蓿的平均发芽天数缩短了0~1 d; 550 μ mol· mol-1处理的苜蓿的平均发芽天数缩短了1 d, 550 μ mol· mol-1处理下苜蓿能以较短时间完成发芽进程。
| 表3 不同CO2浓度下苜蓿各生育期天数 Table 3 Days of each growth period of alfalfa under different CO2 concentrations |
2.2 CO2浓度对苜蓿生育期的影响
研究中CO2浓度升高促进苜蓿生育进程, 生育期有缩短趋势 (表3), 但不同浓度CO2处理或不同苜蓿品种存在差异:其中700 μ mol· mol-1下处理FD1初花期和结荚成熟均提前了4 d; FD6初花期和结荚成熟分别提前了4和5 d; FD11初花期和结荚成熟分别提前了3和6 d。550 μ mol· mol-1处理使FD1初花期和结荚成熟期分别提前了4和3 d; FD6初花期和结荚成熟分别提前了3和4 d; FD11初花期和结荚成熟分别提前了1和4 d。可以看出, CO2浓度越高, 苜蓿生育期越提前。此外, 700 μ mol· mol-1CO2延长了苜蓿的花期, 700 μ mol· mol-1组苜蓿的花期延长了0~1 d。总体来说CO2浓度越高, 生育期越提前; 相同CO2浓度处理下, 秋眠级越高生育期越提前。
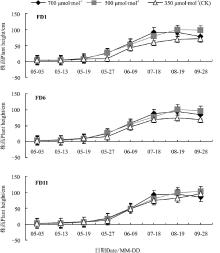
2.3 CO2浓度对苜蓿株高的影响
3种秋眠类型苜蓿在不同CO2浓度下生长状态均呈“ S” 型变化, 即平缓增长-快速增长-停止生长的趋势(图1)。分枝期(6月9日)苜蓿均进入快速生长阶段, CO2浓度越高3种秋眠类型苜蓿的株高越高。盛花期(8月19日)时FD1、FD6株高增长速度相对平缓, 株高达到最大值, 700 μ mol· mol-1组FD1、FD6的株高分别比CK高出32.45%、26.50%; 550 μ mol· mol-1组下FD1、FD6的株高比CK高42.80%、35.54%。
 | 图1 CO2浓度升高对苜蓿株高的影响Fig.1 Effect of elevated CO2 concentrations on plant height of alfalfa |
成熟期(9月28日), 550 μ mol· mol-1组和CK组, FD11的株高直到达到最大值, 株高值分别为102.46、95.76 cm。
5月5日-7月18日, 即处理75 d时, 700 μ mol· mol-1组苜蓿的株高较高, 7月18日-9月28日, 即处理75-150 d时, 550 μ mol· mol-1组苜蓿组仍保持较快的生长势, 而700 μ mol· mol-1组生长减缓。总体来说, 高CO2浓度能促进苜蓿的株高, 且FD11的株高均高于FD1和FD6, 该休眠型苜蓿物质积累更快。
2.4 CO2浓度对苜蓿叶面积的影响
整个生育期期间苜蓿的叶面积都呈“ ∧ ” 型变化, 分枝期至盛花期叶面积逐渐增大, 盛花期至成熟期叶片枯黄缩小, 呈现先升高后降低的趋势。每个生育期, 3种秋眠类型苜蓿的叶面积均在700 μ mol· mol-1处理浓度下达到最大, 其次是550 μ mol· mol-1, CK组最低(表4)。成熟期相同苜蓿品种不同CO2浓度处理下苜蓿的叶面积均大于CK组, 700 μ mol· mol-1处理下, FD1、FD6、FD11的叶面积分别比CK组大45.64%、45.82%、24.53%; 550 μ mol· mol-1处理下, FD1、FD6、FD11的叶面积分别比CK组增大了38.35%、36.22%、7.01%。总体来说, 高CO2浓度处理极显著促进苜蓿叶面积增大(P< 0.01)。除分枝期外, 相同CO2浓度处理下FD11叶面积最大, FD6次之, FD1叶面积最小。
| 表4 CO2浓度升高对苜蓿叶面积(cm2)的影响 Table 4 Effect of elevated CO2 concentrations on leaf area of alfalfa(cm2) |
2.5 CO2浓度对苜蓿茎粗的影响
整个生育期观测结果表明, 除盛花期外, CO2浓度对苜蓿茎粗无显著影响(P> 0.05)(表5)。方差分析结果显示, 分枝期时, 700、550 μ mol· mol-1组内苜蓿的茎粗比CK茎粗分别高出51.94%(P=0.038)、7.00%(P=0.002); 盛花期和成熟期时, 700、550 μ mol· mol-1组内苜蓿的茎粗与CK的茎粗差异不显著(P> 0.05)。对比相同CO2浓度下不同秋眠类型苜蓿的茎粗发现, CK组3种苜蓿的茎粗值差距不大。700 μ mol· mol-1组, 整个生育期FD1的茎粗始终大于FD11茎粗, 但差异不显著(P> 0.05)。550 μ mol· mol-1组, 分枝期, FD1、FD6的茎粗值比FD11分别高出11.93%(P=0.028)、10.55%(P=0.045), 初花期、盛花期和成熟期3个品种茎粗无显著差异(P> 0.05)。总体来说, CO2浓度越高, FD1和FD6苜蓿的茎粗值越大; 高CO2浓度处理下抑制FD11茎粗增加, 促进FD1和FD6的苜蓿茎粗增加。
| 表5 CO2浓度升高对苜蓿茎粗(mm)的影响 Table 5 Effect of elevated CO2 concentrations on stem diameter of alfalfa(mm) |
2.6 CO2浓度对苜蓿产量的影响
CO2浓度越高, 苜蓿的单株鲜草产量越高(图2)。CO2浓度升高对苜蓿单株鲜重有显著影响(P=0.012), 700 μ mol· mol-1 CO2处理下FD1、FD6、FD11的单株鲜重分别高出CK组17.14%(P> 0.05)、30.00%(P< 0.05)、39.64%(P< 0.05)。550 μ mol· mol-1处理下FD1、FD6、FD11的单株鲜重分别高出CK组17.42%(P> 0.05)、13.03%(P> 0.05)、37.94%(P< 0.05)。相同CO2浓度处理下, FD11和FD6苜蓿的单株产量均大于FD1苜蓿, 其中700和550 μ mol· mol-1下FD11苜蓿单株产量显著大于FD1(P=0.024; P=0.032), 而CK组3种秋眠类型苜蓿的单株鲜重差异彼此不显著(P> 0.05)。
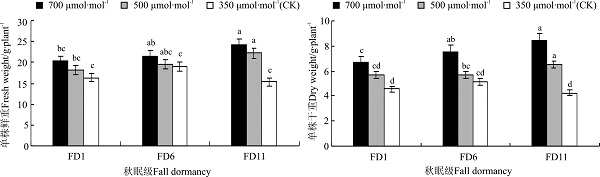 | 图2 CO2浓度升高对苜蓿产量的影响Fig.2 Effect of elevated CO2 concentrations on the yield of alfalfa |
CO2浓度升高显著促进苜蓿的单株干草重(P< 0.05), 且相同苜蓿品种在3个CO2浓度水平处理下单株干重排序均为700 μ mol· mol-1> 550 μ mol· mol-1> CK。其中FD1和FD6在700 μ mol· mol-1处理下单株干重较CK显著增加(P< 0.05), FD11在700和550 μ mol· mol-1处理下单株干重均较CK显著提高(P< 0.05)。相同高CO2浓度处理下, FD1的单株干草重最低, FD6干草重比FD1提高了31.78%~34.54%; FD11干草重均显著大于FD1(P< 0.05)。
3 讨论与结论
3.1 种子发芽对CO2浓度的响应
本研究中高CO2浓度显著促进了3种秋眠类型苜蓿种子的发芽进程, 种子发芽率、发芽势和发芽指数, 发芽天数显著缩短(P< 0.05)。前人研究表明, 白花蝇子草在高CO2浓度下发芽率提高了20%[14]。20% CO2处理可促进蒺藜状苜蓿(Medicago truncatula)种子发芽率, 较CK组发芽率提高2.17倍, 发芽势提高4.14倍, 发芽指数提高3.49倍, 而发芽天数缩短0.73 d[23]。高CO2浓度能使光叶红豆(Adenanthera microsperma)种子萌发率较CK组提高12%[24], 与本研究结果一致。种子发芽率、发芽势和发芽指数的提高均反映了大气CO2浓度对苜蓿种子发芽具有正效应, 适当升高CO2浓度能够促进种子产生乙烯, 而乙烯能够打破种子休眠, 刺激种子发芽[25, 26]。预测的未来大气CO2浓度升高, 植物种子出苗率高, 出苗集中, 趋于一致。发芽指数对CO2浓度变化最为敏感, 可作为未来大气CO2浓度升高情况下苜蓿种植筛选种子的重要参考指标。此外, 高CO2浓度下极非秋眠型苜蓿UC-1465的种子发芽势、发芽率高于秋眠型苜蓿Maverick和半秋眠型苜蓿ABI700。
本研究中700 μ mol· mol-1 CO2浓度对苜蓿的发芽率、发芽指数和平均发芽时间的正效应影响均低于550 μ mol· mol-1 CO2浓度, 说明苜蓿种子发芽有最适CO2浓度范围。这可能是700 μ mol· mol-1 下, O2/CO2降低, 而发芽进行旺盛的呼吸作用需要O2的参与[23, 24, 25, 26, 27]。700 μ mol· mol-1CO2浓度下三叶草(Medicago sativa)、辣椒(Capsicum annuum)、茼蒿(Chrysanthemum coronarium)、康乃馨( Dianthuscaryophyllus)的种子的发芽势等各发芽指数均下降, 其中三叶草的种子活力指数下降了53.1%[27], 进一步证明C3植物种子发芽需要适合的CO2浓度, 最适CO2浓度范围在550~700 μ mol· mol-1。
3.2 植物生长发育对CO2浓度的响应
CO2对植物具有“ 施肥效应” [4]。本研究中CO2浓度升高显著促进苜蓿的株高、叶面积的增加, 最终导致单株鲜重和单株干重显著增加(P< 0.05), 其中苜蓿的单株鲜重平均提高了13.03%~37.64%, 单株干重平均提高了10.65%~98.57%。前人研究表明, CO2浓度提高对紫花苜蓿[28]、夏大豆(Glycine max)[29]、绿豆(Vigna radiata)[8, 30]、飞机草(Chromolaena odorata)[31]和异叶泽兰(Eupatorium heterophyllum)[31]的叶面积、株高、生物量有显著的促进作用。Fischinger等[32]也发现CO2浓度升高, 苜蓿的分枝重, 根系重以及总物质量均显著增加, 根瘤物质量增加, 保证了高CO2浓度下固氮能力的提升。CO2浓度920 μ mol· mol-1时促进马铃薯(Solanum tuberosum)的株高、茎粗、叶面积和块茎生长, 而上升至3 600 μ mol· mol-1时则明显抑制马铃薯的生长发育[33]。可以发现, CO2浓度对植株的形态特征的影响因植株的种类和CO2的浓度而异。结合前人研究成果, 本研究证明了在未来大气CO2浓度升高的趋势下, 豆科植物产量将大幅度增加[28, 29, 30]。
本研究中CO2浓度对茎粗影响不显著(P> 0.05), 高CO2浓度促进秋眠型苜蓿Maverick和半秋眠型苜蓿ABI700的茎粗增加, 而对极非秋眠型苜蓿UC-1465的茎粗无影响, 与Sun等[33]和Wang等[34]研究结果不完全一致, Wang等[34]认为CO2浓度升高能增加豆科植物的固氮能力, 茎基部直径也增加。高CO2浓度促进了3类秋眠苜蓿的生育期提前了3.25~5.75 d。同样CO2浓度的倍增大大促进了小麦生育期提前[35]。CO2浓度升高缩短植物生育期, 叶片提前衰老, 是对光合适应现象的另一种合理解释[36]。此外, 高CO2浓度处理后不同秋眠型苜蓿品种生育期存在差异, 这可能与供试品种生育期的长短有关:高浓度CO2环境下苜蓿生育期短的品种(FD11)和较长生育期品种((FD1和FD6)的生长优势得到进一步加强和促进, 从而导致生育期缩短更为明显, 但这一假设尚需更多品种试验的证实。综合而言, CO2浓度升高加速了苜蓿的种子发芽和生长代谢, 使光合产物更多地向碳“ 汇” 器官(叶和茎) 运输, 以用于生长[37]。
因此, 在一定浓度范围内, 苜蓿种子的发芽率、发芽指数、发芽势、株高、叶面积、生物量随着CO2浓度升高而升高。550、700 μ mol· mol-1CO2浓度处理显著促进发芽率、发芽势、发芽指数, 缩短平均发芽天数。其中550 μ mol· mol-1浓度处理下苜蓿种子的发芽率、发芽势最高, 不同品种间, ABI700和UC-1465种子发芽率和发芽势更高。高CO2浓度使得3种秋眠类型苜蓿生育期提前3~6 d, 且CO2浓度越高生育期越提前。75 d高CO2浓度处理后UC-1465的生长发育速度进程加快, 比Maverick和ABI700提前1~2 d成熟。高CO2浓度能显著促进3种秋眠类型苜蓿株高、叶面积和单株干重增加, 但对苜蓿茎粗无显著影响。
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|