

{kind=link}
{kind=link}
播种量对高寒牧区林纳燕麦生产性能及光合特性的影响
[肖雪君1 , 周青平2  , 陈有军
, 陈有军2 , 杜忠2 , 柏晓玲1 , 田莉华2 , 彭先琴3 ]
, 陈有军|
|


第一作者:肖雪君(1992-),女,四川宜宾人,硕士,主要从事牧草栽培育种研究。E-mail:[email protected]
播种量是保障牧草高产质优的一个重要方面,合理的播种量除取决于牧草品种自身的生物学特性外,还受到外界条件的影响。为探明适合川西北高寒牧区自然气候条件下林纳燕麦( Avena sativa cv. LENA)的最适播种量,本研究设置了120(A)、165(B)、210(C)、255(D)、300(E)和345 kg·hm-2(F)共6个播种量,分析播种量对林纳燕麦生产性能及光合特性的影响。结果表明,随播种量的增加,林纳燕麦物候期表现出了提前的趋势,在出苗期不明显,但在其它生育期较明显;播种量对燕麦的株高无显著影响( P>0.05),但在大多数的取样期,林纳燕麦的株高随着播种量增加表现为减小的趋势;随着播种量增加,茎粗呈下降趋势,尤其在生育期后期呈显著降低趋势( P<0.05);播种量对茎叶比无显著影响( P>0.05),但随着生育期进程,茎叶比呈显著降低趋势( P<0.05);在高播种量(E、F)下,鲜草产量和干草产量均表现为增大趋势,产草效果最佳;在开花期,低播种量(A、B、C)的气孔较活跃,而高播种量(E、F)胞间CO2浓度较低。在该研究区域,在行距30 cm,播种量300或345 kg·hm-2下,林纳燕麦的生产性能及光合特性最理想,产草效果最佳。本研究结果可为川西北高寒牧区林纳燕麦的高产栽培及推广应用提供理论支持。
, Chen You-junSeeding rate is an important factor to guarantee high yield and quality of forage grass. Besides the biological characteristics of the forage grass, the reasonable seeding rate may also be affected by external conditions. In order to determine the optimum seeding rate in northwest alpine pastoral regions of Sichuan, Avena sativa cv. LENA was chosen as the material to explore the effects of six seeding rates (A: 120 kg·ha-1, B: 165 kg·ha-1, C: 210 kg·ha-1, D: 255 kg·ha-1, E: 300 kg·ha-1, and F: 345 kg·ha-1) on production performance and photosynthetic characteristics. Phenophase of A. sativa cv. LENA decreased with increasing seeding rates, and at the seedling stage, there was no significant difference, but at other stages, the differences were obvious. For plant height, there were no significant differences among the six seeding rates. The thickness of stems decreased significantly with the increasing seeding rates, and the tendency was more obvious at the latter stages. The stem-to-leaf ratio showed no significant differences between different seeding rates ( P>0.05), but the ratio decreased significantly with the growth stages ( P<0.05). The fresh and dry hay yield increased significantly with the growth stages, and the largest dry hay yield was at E and F seeding rates. At the flowering stage, the stoma was more active at the A, B, and C seeding rates, and the intercellular CO2 concentration was lower at E and F seedling rate. Under the conditions of 30 cm row spacing and 300 kg·ha-1 or 345 kg·ha-1 seeding rate, A. sativa cv. LENA production performance and photosynthetic characteristics were ideal. These results provide theoretical support for A. sativa cv. LENA cultivation practices in northwest alpine pastoral regions of Sichuan to attain higher yields.
燕麦(Avena sativa)为一年生禾本科草本植物, 在我国种植时间较长, 主要分布在东北、华北和青藏高原高寒地区[1]; 从类型上主要分为带稃型和裸粒型[2, 3]。燕麦是重要的粮饲兼用作物[2], 具有草产量高、适口性好、营养丰富等特点[4, 5], 是重要的圈窝种植牧草, 可有效缓解家畜“ 冬瘦、春乏” 的现象, 在维系畜牧业的发展上具有其它牧草不可替代的作用, 在高寒牧区倍受广大牧民的青睐[2]。青藏高原具有特殊的自然环境条件, 无法满足大多数作物正常生长需要, 而燕麦由于喜冷凉、耐贫瘠、耐盐碱、抗旱、耐寒, 在青藏高原高寒牧区得到大面积的推广及应用[5, 6], 是高寒草地畜牧业稳定发展中必不可少的饲草作物[7, 8, 9]。
播种量是保障牧草高产优质的一个重要方面, 采用合理的播种量除取决于牧草自身的生物学特性外, 还受外界条件的影响, 如土壤条件、气候条件等的影响。目前, 对燕麦的研究工作主要集中在栽培技术[10, 11]、播种期[12]、物候期[13]、营养价值[14]、抗性[15, 16]、品比试验[17, 18]及种质资源的遗传多样性[19, 20]等方面, 为燕麦优质品种的筛选以及推广应用提供理论依据, 在维系生态草地畜牧业的可持续发展中起着至关重要的作用。随着燕麦品种的不断增加, 播种量的研究还没有实现良种良法, 比如早熟品种与晚熟品种在不同地区的适宜播种量。青藏高原区域由于地区间水热条件差异较大, 栽培品种及栽培方法也明显不同, 燕麦单播适宜播种量的选择一直没有普适的结论。而且, 前期的研究[11, 17]也表明, 合理的播种量会影响牧草的群体数量和结构, 缓和群体与个体之间生存资源竞争的矛盾, 有利于牧草干物质积累, 从而达到牧草的高产。“ 青藏高原特有草种质资源创新及草地生态恢复” 研究团队主要从事抗逆牧草的种质挖掘与创新, 通过前期开展的室内抗性试验[16]及室外品比试验[17]得出, 林纳燕麦的抗性比较好, 适宜在青藏高原高寒牧区推广种植。因此, 为了探明林纳燕麦在川西北高寒牧区自然条件下的最适宜播种量, 于2015年5月至10月在四川省红原县西南民族大学青藏高原基地, 开展不同播种量对林纳燕麦生产性能及光合特性影响的试验研究, 通过对比分析林纳燕麦在不同播种量处理下其生产性能及光合特性的差异性, 筛选出适合当地自然环境条件下的最优播种量, 以期为川西北高寒牧区林纳燕麦的高产栽培及推广应用提供理论支持。
研究区位于青藏高原东缘四川省阿坝州红原县境内(31° 50'-33° 22' N, 101° 51'-103° 23' E)的西南民族大学青藏高原基地, 平均海拔3 500 m。气候属于大陆高原季风气候, 年降水量650~800 mm, 80%集中于每年的5月-9月[21]。种植区域土层深达40 cm, 土壤有机碳含量较高, 为215~280 g· kg-1, 总氮和磷含量较低, 总氮含量为6~10 g· kg-1, 有效磷含量为3~7 mg· kg-1, pH 4.6左右[22]。
本研究共设置6个播种量, 分别为120、165、210、255、300和345 kg· hm-2, 对应的有效株数分别为385万、529万、673万、818万、962万、110 6万株· hm-2。试验采用完全随机区组设计, 每个播种量设3个重复, 每个重复1个小区, 共计18个试验小区, 小区面积为5 m× 3 m。种植前所有试验小区机耕翻耙, 耕深为25 cm, 然后人工整地、耙平、划区播种。于2015年5月28日统一播种, 采用开沟条播, 播深3~5 cm, 行距30 cm。整个试验期间无灌溉和施肥。在播种后第15天进行人工除草一次。
1.4.1 物候期 采用整体观察法, 观测并记录每个小区的物候, 以试验小区内植株的50%进入某物候期为基准。
1.4.2 茎粗和株高 燕麦出苗后, 在每个小区远离边行30 cm处, 随机选取5株挂牌定株, 用游标卡尺测定其茎粗, 用钢卷尺测定其株高, 每10 d测定一次。
1.4.3 产量性状 在每个小区远离边行30 cm处随机取1 m样段, 齐地刈割, 茎、叶(小穗跟叶分到一起)分开分别称其鲜重后, 放入烘箱(65 ℃)烘干至恒重后, 再分别称其干重, 测其鲜草产量、干草产量、鲜干比及茎叶比。茎叶比(stem/leaf, S/L)是在各时期所取得的茎与叶生物量的比值, 其中:鲜茎叶比为SF/LF, 干茎叶比SD/LD。每个小区取样1次, 每个播种量处理为3个重复。
1.4.4 相对生长速率 相对生长速率(relative growth rate, RGR)是在单位时间内生物量的净积累。利用各时期取得的干草产量数据, 计算相对生长速率, 计算公式:
RGR=
式中:RGR为平均相对生长率[g· (g· d)-1], W2和W1分别为t2和t1时所获取的燕麦的生物量(g· m-2)。
本研究使用SPSS 19.0(SPSS Inc., version19.0)软件对所测指标进行单因素方差分析, 然后用Duncan法对测定指标进行多重比较, 分析不同播种量之间的差异性, 最后用Excel 2010软件制作图表。
2.1.1 播种量对林纳燕麦物候期的影响 不同播种量下林纳燕麦物候期存在差异性(表1)。高播种量下出苗期较早, 随着播种量的增加, 物候期越来越短, 各个发育时期都呈现提前趋势。低播种量(120、165、210和255 kg· hm-2)在10月下旬全部完成生育期, 而高播种量(300和345 kg· hm-2)则在10月上旬完成生育期, 播种量为120 kg· hm-2时, 生育时长为145 d, 345 kg· hm-2时为137 d, 相差8 d。综合来看, 在生育期的前期, 不同播种量间的生育时差间隔较小, 在抽穗期生育时差间隔最大(9 d), 之后保持不变。结果表明, 高播种量可以使燕麦物候期提前, 且在中后生育期播种量的作用会更加明显。
2.1.2 播种量对林纳燕麦株高的影响 随着生育期的进程, 林纳燕麦的株高不断增加(表2)。在大多数的取样期, 林纳燕麦株高随着播种量增加呈减小的趋势, 除6月20日和10月17日不同播种量对株高的影响差异显著外(P< 0.05), 其它取样期内播种量对株高的影响均不显著。完熟期(10月17日)低播种量(120 kg· hm-2)下的株高显著高于高播种量(255 kg· hm-2)下的株高。
| 表1 播种量对林纳燕麦物候期及生育时长的影响 Table 1 Effect of seeding rate on phenology and the whole growth duration of Avena sativa cv. LENA |
| 表2 播种量对林纳燕麦株高(cm)的影响 Table 2 Effect of seeding rate on the height(cm) of Avena sativa cv. LENA |
2.1.3 播种量对林纳燕麦茎粗的影响 从整个生育周期来看, 播种量对燕麦的茎粗存在影响(图1)。在生育期前期(7月1日-7月30日), 播种量对燕麦茎粗无显著影响(P> 0.05), 但是在生育后期播种量显著影响燕麦茎粗(P< 0.05)。另外, 不同播种量处理下, 林纳燕麦茎粗并不是持续上升的, 其中7月30日到8月20日的下降趋势, 可能是研究区域7月30日之前降水量丰富, 但是之后降水量少, 气候干旱, 茎秆含水量降低所致。整体看来, 低播种量下, 茎粗较大, 在高播种量(300、345 kg· hm-2)下, 茎粗较小。
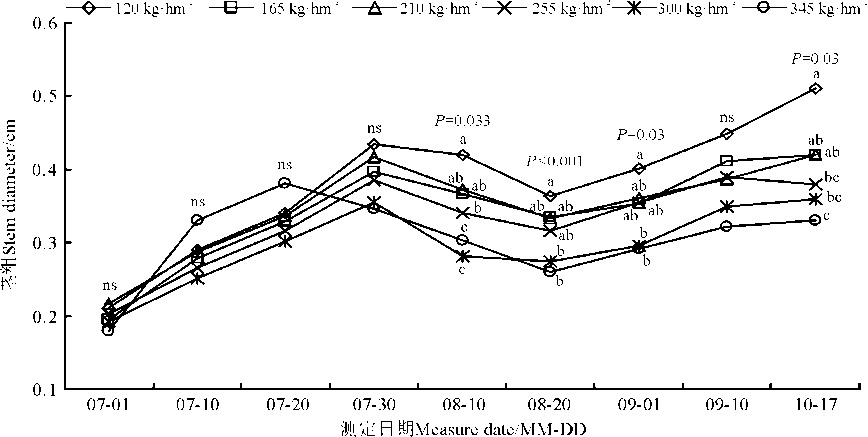 | 图1 播种量对林纳燕麦茎粗的影响Fig. 1 Effect of seeding rate on stem diameter of Avena sativa cv. LENA |
播种量对燕麦不同发育期茎叶比的影响明显不同(表3)。在抽穗期和完熟期, 不同播种量下, 燕麦茎叶比(鲜、干)大小不同, 但是无显著差异(P> 0.05)。在拔节期, 播种量仅对干茎叶比有显著影响(P< 0.05)。
| 表3 播种量对林纳燕麦不同发育期茎叶比(鲜、干)的影响 Table 3 Effect of seeding rate on the stem to leaf ratio of Avena sativa cv. LENA at different growth stages |
在开花期, 播种量对鲜、干茎叶比均有显著影响(P< 0.05), 茎叶比随着播种量的增加而增加。综合整个生育周期来看, 不同播种量下茎叶比(鲜、干)随着生育期的进程而增加, 且均在完熟期达到最大值。
林纳燕麦鲜草产量随着生育期进程逐渐增加(表4), 但是其生长速率则表现出随生育期进程逐渐降低(表5)。在拔节期, 播种量对林纳燕麦鲜草产量有显著影响(P< 0.001), 最大值出现在播种量为255 kg· hm-2时(5 936 kg· hm-2); 在抽穗期和完熟期, 鲜草产量整体上表现为随播种量的增加而增加的趋势, 在播种量为345 kg· hm-2时达到最大(P< 0.05)。生长速率在拔节期到抽穗期较大, 但不同播种著差异(P> 0.05), 说明6个播种量在拔节期都生长较快; 抽穗期到开花期不同播种量间生长速率存在显著差异(P=0.011), 其中播种量为345 kg· hm-2时的生长速率显著小于120、165和210 kg· hm-2时(P< 0.05); 在开花期到完熟期, 生长速率较小, 但播种量对其仍然有显著影响(P=0.028), 且在播种量为345 kg· hm-2时生长速率最大。
| 表4 播种量对林纳燕麦不同发育期鲜草产量的影响 Table 4 Effect of seeding rate on fresh yield of Avena sativa cv. LENA at different growth stages |
| 表5 播种量对林纳燕麦不同发育期生长速率的影响 Table 5 Effect of seeding rate on the relative growth rate of Avena sativa cv. LENA at different growth stages |
不同播种量处理下, 干草产量均随生育期进程而增加; 播种量对不同生育期的干草产量均有显著影响(P< 0.05)(表6)。在拔节期, 随着播种量的增加干草产量呈现增加的趋势, 在255和345 kg· hm-2的播种量下的干草产量显著大于其它播种量(P< 0.05), 255 kg· hm-2时为1 978.7 kg· hm-2, 345 kg· hm-2时为1 726 kg· hm-2; 在抽穗期, 345 kg· hm-2播种量处理下草产量达到最大值, 为4 889.7 kg· hm-2, 且显著大于120、165和210 kg· hm-2播种量处理(P< 0.05); 在开花期, 干草产量在不同播种量间表现为300> 210> 345> 120> 255> 165 kg· hm-2, 且210、300和345 kg· hm-2播种量处理下显著大于120、165和255 kg· hm-2播种量处理(P< 0.05); 在完熟期, 播种量为345 kg· hm-2时的干草产量显著大于播种量为165、255和300 kg· hm-2播种量处理(P< 0.05)。综合整个生育期可知, 高播种量下的干草产量显著高于低播种量的干草产量, 表明适当增加燕麦的播种量可以提高干草产量。
| 表6 播种量对林纳燕麦不同生育期干草产量的影响 Table 6 Effect of seeding rate on hay yield of Avena sativa cv. LENA at different growth stages |
2.5.1 播种量对林纳燕麦不同生育期叶绿素相对含量的影响 林纳燕麦叶绿素相对含量在不同播种量下存在差异, 但是在整个生育期内不同的播种量间呈现相似的变化规律(图2)。在7月1日到7月10日, 叶绿素相对含量迅速增加; 7月10日到9月10日, 叶绿素相对含量变化比较缓慢; 9月10日以后, 叶绿素相对含量迅速下降。由于播种量明显影响燕麦的物候期, 所以不同播种量燕麦的叶绿素相对含量在不同时期间达到最大值, 120 kg· hm-2播种量在7月30日时达到最大, 210 kg· hm-2播种量在7月10日达到最大, 其余播种量更早, 在7月1日就达到最大。综合整个生育期可知, 燕麦叶绿素相对含量都随着播种量的增加呈现减小的趋势, 尤其是在生育期的前期(7月1日)和后期(10月17日), 不同播种量间存在显著差异(P< 0.05)。
 | 图2 播种量对林纳燕麦不同生育期叶绿素相对含量的影响Fig. 2 Effect of seeding rate on relative chlorophyll content of Avena sativa cv. LENA at different growth stages |
2.5.2 播种量对林纳燕麦开花期光合特性的影响 开花期是植物生长的重要时期, 播种量显著影响了林纳燕麦开花期的光合特性(P< 0.05)(表7)。林纳燕麦净光合速率、气孔导度、胞间CO2浓度均随着播种量增加而降低, 表明播种量的增加导致植物个体之间竞争增强。但是, 播种量对林纳燕麦的蒸腾速率的影响不显著(P> 0.05)。
| 表7 播种量对林纳燕麦开花期光合特性的影响 Table 7 Effect of seeding rate on photosynthetic capacity of Avena sativa cv. LENA at the flowering stage |
自然界中, 植物个体通过改变自身的形态特征, 影响其对环境资源的利用[23], 形成与外界环境相适应的生存策略, 是在特定生境条件下长期适应的结果。密度制约在植物种群中普遍存在, 是种群极其重要的选择压力[24], 本质是因为自然界中存在着有限的资源(光资源、生存空间、水分和养分), 密度制约主要是种群内植株个体对有限资源的竞争, 通过竞争改变种群内植株的各性状参数[25], 实现其最适宜、最有利的生存和繁殖状态。
描述牧草生长状况最合适的指标就是株高, 同时株高也是反映牧草产量的一个特征值[26]。株高与产量、覆盖度呈正相关关系[27, 28], 是能否获得较高产量的保证[29], 同时也是评价牧草经济产量形成以及衡量其生长状况较为理想的一个特征值[27], 是反映草地生产能力的重要生产指标[30]。整体而言, 播种量对出苗后生长期内燕麦株高的影响较小, 仅出苗期和完熟期的株高差异较为显著。表明同一燕麦品种营养生长阶段及生殖生长前期的株高是一个相对稳定的性状, 播种量对其影响较小, 但在生殖生长后期(完熟期), 低播种量可以减小种间竞争, 提高燕麦株高。一般植株高度是由生物学特性决定[31], 但环境因素也会对株高产生影响, 在播种量低时, 因为植株个体间营养竞争较弱, 植株能够获得更多的养分和能量, 从而生长较快; 但是随着播种量的增大, 植株间对资源的竞争变得更加激烈, 在资源有限的情况下, 种群内个体可获得的资源量减少, 从而引起株高的响应变化[32]。本研究结果与前期在燕麦上开展的类似研究所得结果[33, 34]一致。
茎粗也是影响牧草产量的主要原因之一, 也是决定其利用方式的重要根据[35]。本研究发现, 茎粗在生长初期各播种量处理间差异不大, 随着播种量增加呈下降趋势, 而且这种趋势在生育期后期较显著。这是因为, 生长初期土壤养分含量较高, 加之各处理间光资源竞争较小, 而随着生长进程, 在气候条件较差的干旱时期, 为了适应环境的改变, 茎秆含水量降低, 茎粗变小。
在饲草生产中, 地上生物量定义为单位面积内饲草的积累量, 可以用鲜重或干重计算[36], 生物量可体现生产力并反映其经济效益[7]。对于燕麦来说, 判断其饲用价值高低的重要指标之一就是产草量, 主要受光照、温度、水分等环境因素的影响, 而且也是株高、茎粗、生长速率等性状指标的综合体现[37]。本研究表明, 林纳燕麦的鲜草产量和干草产量均表现为在高播种量下较高的趋势, 在300、345 kg· hm-2播种量下产草效果最佳, 该结果与对皖麦44(Triticum aestivum)[38]的研究结果大致相同, 表明适当增加燕麦的播种量可以提高养分利用效率, 提高产草量。研究表明, 作物干物质的积累存在着明显的播种量效应, 随着播种量的增大, 整个群体的干物质积累量也增加[39, 40]。但是, 当超过最佳播种量时, 由于个体之间存在着对资源的竞争, 导致干物质积累能力降低, 势必会引起干物质积累量的下降。同样, 当牧草播量过低时, 由于作物群体密度小, 养分利用效率下降, 减少了作物干物质的积累量。综合整个生育期可知, 在开花期, 300 kg· hm-2播种量的产草量高于345 kg· hm-2播种量的产草量, 差异不明显; 在完熟期, 345 kg· hm-2播种量的草产量显著高于其它播种量(P< 0.05)。虽然随着播种量的增大, 燕麦个体之间存在着对水分、养分的竞争, 但是个体间的资源竞争还未引起群体干物质积累能力的下降, 如果进一步增大燕麦的播种量, 可能群体产草量会增加, 但是也可能已经达到最大播种量, 这需要后期开展进一步的播种量和养分竞争的相关研究。
茎叶比的大小可以衡量牧草经济性状的高低, 也能反映其适口性和干草品质。本研究表明, 茎叶比随着生育期延长而增加, 因为随着燕麦植株高度的不断生长, 茎秆积累量明显增多, 比值逐渐增大, 主要是由燕麦的生长特性所决定的。但是, 在燕麦的不同生育期(开花期除外), 茎叶比在不同播种量间差异不显著, 总体上呈现增加的趋势, 表明随着播种量的增大, 燕麦个体间对资源的竞争越来明显。
光合作用主要是增加作物干物质的积累[41, 42], 是作物生产力形成与演化的基础, 但是光合作用受多种因素的影响(土壤水分、光照、温度以及植物本身特性)[43], 因此, 在不同时期, 光合特性也存在差异。本研究发现, 开花期的气孔导度在播种量120 kg· hm-2时显著高于300和345 kg· hm-2播种量时, 165 kg· hm-2播种量显著高于300 kg· hm-2, 210 kg· hm-2播种量显著高于300和345 kg· hm-2播种量。说明播种量120、165和210 kg· hm-2的气孔较活跃, 而300 kg· hm-2由于种间竞争较强导致胞间CO2浓度较低。而有研究表明, 气孔导度与蒸腾速率、胞间CO2浓度关系十分密切[43]。开花期是燕麦生长的一个重要时期, 研究该时期的光合特性对燕麦栽培研究具有重要的意义。不同播种量处理, 其密度存在差异, 导致植株光合特性也产生差异。因此, 研究不同生育期的光合特性对建立高产、高品质的燕麦草生产将是今后研究的重点及方向。
林纳燕麦的适应性强, 饲用价值优良, 是一种非常优良的草饲兼用作物, 对高寒牧区草地畜牧业的可持续发展具有重要的战略意义。然而由于青藏高原自然条件的限制, 目前适于高寒牧区林纳燕麦的栽培技术还比较缺乏。通过开展的林纳播种量的研究, 主要得到以下几个结论:
1)在该研究区域, 林纳燕麦的生产性能及光合特性显著受到播种量的影响。总体上表现为:在适当的播种量范围内, 随着播种量的增加, 林纳燕麦的生育期提前、产草量增加。对于积温低、生长季短的高寒牧区来说, 这都是非常有利的。因此, 适当增大林纳燕麦的播种量, 可以提高养分利用效率, 收获较高的饲草产量。
2)综合林纳燕麦的各项生产性能(株高, 茎粗, 茎叶比, 干草产量)及光合特性(净光合速率、气孔导度、胞间CO2浓度、叶绿素含量)来看, 在该研究区域, 行距30 cm, 播种量300或345 kg· hm-2下, 林纳燕麦的表现最佳, 适宜在川西北高寒牧区推广。
3)植物的光合作用是生产力形成与演化的基础。目前, 国内外关于植物光合特性与生长特性相关性的研究大部分集中于小麦(Triticum aestivum)作物, 对青藏高原高寒牧区种植燕麦的研究就更少了, 严重限制了对燕麦能够适应高寒区域复杂环境的适应性机理的认知, 以及对燕麦饲草高产、稳产的研究。因此, 研究燕麦不同生育期的光合特性对建立高产、高品质的燕麦栽培草地具有重要的理论和现实意义。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
播种量对高寒牧区林纳燕麦生产性能及光合特性的影响
肖雪君, 陈有军
Effect of seeding rate on production performance and photosynthetic characteristics of Avena sativa cv. LENA in alpine pastoral regions
Xiao Xue-jun, Chen You-jun