
植物内整流K+通道AKT1的研究进展
[胡静1  , 胡小柯
, 胡小柯1 , 尉秋实1 , 袁惠君2 ]
, 胡小柯|
|


K+是植物生长发育所必需的大量营养元素。内整流K+通道( Arabidopsis K+ transporter 1,AKT1)属于Shaker家族,是介导K+吸收的重要通道,为质膜的K+感应器,参与调节细胞的生长发育、调控气孔运动及植物蒸腾作用,能够提高植株的抗旱耐盐性,因而在植物生长过程中具有重要作用。该文概述了AKT1的结构、组织表达定位和表达调控及功能等方面的研究进展,并提出采用蛋白组学、基因工程技术及RNAi手段深入研究K+、Na+吸收及转运的协同调控机制,提高作物对土壤中K+的利用效率及AKT1在植物生理代谢、抗逆性中的作用。
Potassium (K+) is one of the major nutrients, essential for plant growth and development. Inward rectifying K+ channel AKT1 belongs to Shaker-type K+ channels, and a K+ sensor in the plasma membrane has been shown to play an important role in mediating K+ uptake. AKT1 is also essential for normal growth and development, stomatal action and transpiration, and improving the drought and salt tolerance of plants. In this review, we consider recent advances in molecular structure, localization, expression, regulation and function of AKT1. Finally, research directions for the future are proposed, including the use of proteomics, gene engineering technology, and posttranscriptional gene silencing, in order to further investigate the role of AKT1 in plant physiological metabolism.
土壤中K+的浓度大部分在0.025~5 mmol· L-1范围内波动[1, 2], 而细胞质中的K+含量却高至60~200 mmol·
近年来, 有关内整流K+通道AKT1的研究受到学术界的广泛关注, 随着研究的不断深入, 发现该K+通道的功能不仅仅局限于介导K+吸收, 为此, 本文就内整流K+通道AKT1的结构、组织表达定位及功能方面的研究进展进行概述, 在此基础上提出采用蛋白组学、基因工程技术及RNAi手段深入研究K+、Na+吸收及转运的协同调控机制, 以期提高作物对土壤中K+的利用效率及AKT1在植物生理代谢、抗逆性中的作用。
1992年, Sentenac等[21]通过酵母钾亏缺突变体互补法从拟南芥中首次分离了K+通道AKT1, 该基因含11个外显子及10个内含子, 位于第2条染色体上, 是第1个克隆到的植物内向整流K+通道基因。同时分离出来的还有K+通道KAT1。继拟南芥后, 先后在番茄(Lycopersicon esculentum)[17]、玉米(Zea mays)[22]、胡萝卜(Daucus carota)[23]、水稻(Oryza sativa)[24]、大麦(Hordeum vulgare)[25]、小麦(Triticum aestivum)[26]、小花碱茅(Puainellia tenuiflora)[27]、霸王(Zygophyllum xanthoxylum)[28]、陆地棉(Gossypium hirsutum)[29]及盐地碱蓬(Suaeda salsa)[30]等中克隆得到AKT1(表1)。可见, AKT1已引起了研究者的广泛关注。
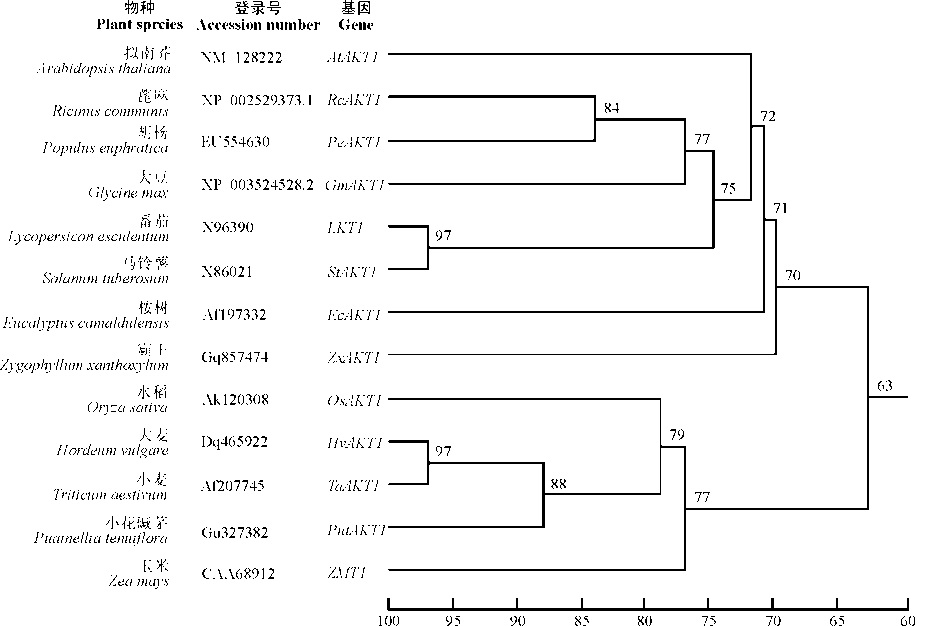
如表1所示, 高等植物中内整流K+通道AKT1基因约含2 570~2 810个核苷酸的开放阅读框(open reading frame, ORF), 编码857~935个氨基酸。对13种植物的内整流K+通道AKT1的氨基酸序列进行了分析(图1), 发现单子叶植物之间的同源性为76%~97%, 双子叶植物之间的同源性为68%~97%, 而单子叶与双子叶植物之间的同源性仅为63%~68%。可见, 不同植物的内整流K+通道AKT1具有一定的差异, 特别是单子叶与双子叶植物之间, 可能是因为它们在长期的进化过程中产生了分异。
| 物种 Plant species | 基因名称 Gene names | 基因库登陆号 GenBank accession numbers | 氨基酸残基 Amino acids | 参考文献 References |
|---|---|---|---|---|
| 拟南芥Arabidopsis thaliana | AKT1 | NM_128222 | 857 | [21] |
| 番茄Lycopersicon esculentum | LKT1 | X96390 | 883 | [31] |
| 玉米Zea mays | ZMK1 | CAA68912 | 887 | [22] |
| 胡萝卜Daucus carota | DcAKT1 | AJ697979 | 873 | [23] |
| 葡萄Vitis vinifera | VvAKT1 | XP_002281787.1 | 872 | - |
| 水稻Oryza sativa | OsAKT1 | AK120308 | 935 | [24] |
| 大麦Hordeum vulgare | HvAKT1 | DQ465922 | 898 | [25] |
| 小麦Triticum aestivum | TaAKT1 | AF207745 | 897 | [26] |
| 马铃薯Solanum tuberosum | SKT1 | X86021 | 883 | [32] |
| 烟草Nicotiana tabacum | NtAKT1 | AB196790 | 933 | - |
| 蓖麻Ricinus communis | RcAKT1 | XP_002529373.1 | 901 | - |
| 桉树Eucalyptus camaldulensis | EcAKT1 | AF197332 | 873 | - |
| 胡杨Populus euphratica | PeAKT1 | EU554630 | 877 | - |
| 小花碱茅Puainellia tenuif lora | PutAKT1 | GU327382 | 900 | [27] |
| 霸王Zygophyllum xanthoxylum | ZxAKT1 | GQ857474 | 869 | - |
| 陆地棉Gossypium hirsutum | GhAKT1 | KF294166 | 875 | [29] |
| 盐地碱蓬Suaeda salsa | SsAKT1 | - | 879 | [30] |
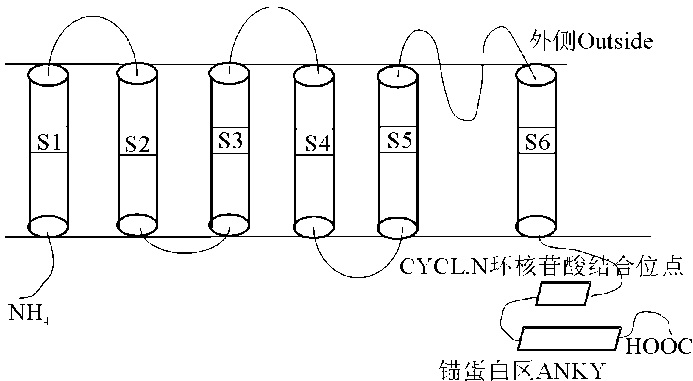
植物中的Shaker通道是目前研究最为清楚的K+转运家族, 研究表明, 在序列及结构上, 其与动物中Shaker家族的K+通道相似[33]。内整流K+通道为典型的Shaker家族成员, 其具有6个跨膜区(S1~S6), S4是通道的电压敏感区, 是一个具有Arg/Lys-Xaa-Xaa-Arg/Lys重复序列区, S5~S6间是离子通道孔径形成区, 中间在内, 两端暴露在外, 为β 发夹形状, 且有一个孔道区域-P环(pore, P), 此区域在K+通道中非常保守, 并含有具有高选择性K+通道特点的基本序列-GYGD/E[21, 34]。N端为胞质内一个很短的结构域(大约60个氨基酸); C端也同样位于胞质内, 具环核苷酸结合位点(CYCL.N.:cyclic nucleotide-binding site)和一个在大部分K+通道中存在的锚蛋白区(ANKY:ankyrin-related domain)[35](图2)。锚蛋白可使蛋白与细胞骨架结合, 或促进蛋白与蛋白之间的相互作用[36]。已有研究认为对Ca2+敏感的CBL(calcineurin B-like Ca2+)和CIPK(CBL-interacting protein kinase)家族同2C型蛋白磷酸酶可以形成一个蛋白磷酸化或去磷酸化的网络从而来调节植物AKT1类K+通道, 其中CIPK就是与AKT1的锚蛋白区域进行相互作用的[37, 38]。此外, 锚蛋白区是区分AKT1类与KAT1类基因的标志, 前者有锚蛋白区, 而后者没有[39, 40]。
Basset等[41]报道AKT1主要在拟南芥成熟根表皮和根毛中表达。Lagarde等[18]以AKT1的cDNA为探针, 通过Northern印迹杂交检测AKT1主要在拟南芥成熟根中表达, 而在叶组织中表达量非常少; 利用Western印迹杂交对油菜(Brassica napus)根系进行定位表达, 证实AKT1基因的表达集中在根的细胞质膜上; 利用AKT1-GUS融合基因检测, 进一步表明了AKT1主要在拟南芥成熟根质膜细胞中表达。而番茄中的LKT1则主要分布在根毛[31]。PutAKT1-GFP融合蛋白轰击洋葱(Allium cepa)表面, 瞬间表达表明, PutAKT1主要定位在细胞质膜上[27]。水稻的OsAKT1主要在根和胚芽鞘中表达, 其中在表皮和内皮层细胞中的表达非常强烈, 而相比之下, 脉管及外皮层等部位的表达量则很低; 叶中也有少量的表达[24, 42]。SKT1定位在马铃薯(Solanum tuberosum)根中[32]。HvAKT1也主要在大麦的根中表达, 叶组织中也有较低的表达量[25]。而GhAKT1却主要在陆地棉叶中表达, 根及茎中的表达量较低, 但也定位在细胞质膜上[27]。综上, 在高等植物中, AKT1主要在植物根的细胞质膜中表达, 可能参与K+吸收。
植物肿瘤细胞在增殖过程中受AKT1介导的K+吸收的促进, 且在AKT1敲除的突变体中, 肿瘤细胞的发育则明显受到抑制[43]。经农杆菌侵染引起肿瘤的叶组织与未经侵染中的K+通道基因的表达谱进行了对比分析, 结果发现二者中TPK表达丰度没有明显变化, 与未侵染的相比, 经农杆菌侵染后的叶组织中AKT1表达量却显著增加, AKT2和GORK转录丰度则显著下降[43, 44]。由此可见, K+通道与植物细胞的生长发育有关。与拟南芥野生型相比, akt1突变体植株在含有100 μ mol· L-1或更低浓度的K+的介质中生长受到了抑制、吸收86Rb+的量减少, 同时检测不到K+内流[17]。当AKT1突变体幼苗置于外界低K+浓度环境时, 与野生型相比, 其根毛很短[27]。因此, AKT1在调节植物生长发育方面起着非常重要的作用。
内整流K+通道AKT1主要在根中表达, 说明其可能参与K+吸收[24, 25, 27, 42]。对akt1突变体的研究表明, AKT1在低浓度K+(10 μ mol· L-1)条件下介导植物根细胞K+吸收[17]。通过电压钳与膜片钳技术发现CIPK23和CBL1或CBL9协同调节可以激活AKT1, 并进一步证明AKT1在植物吸收K+方面起着非常重要作用, 特别是在外界高K+条件下[38]。在K+饥饿条件下, 小麦根部的TaAKT1的表达丰度显著上调, 进一步通过膜片钳分析, 发现小麦根中[K+]in电流受K+饥饿的刺激, 可见TaAKT1介导根中K+的内向电流[26]。通过吸收动力学对突变体athak5和akt1-1的研究表明, AtHAK5介导低浓度K+下K+吸收, AKT1在高浓度范围内(0.9 mmol· L-1)介导K+的吸收[16], AtHAK5与AKT1在K+吸收中所起的作用如下:植物根细胞质膜中存在着负责K+吸收的两个主要系统, 高亲和性和低亲和性系统, 前者即系统Ⅰ 在外部K+浓度较低时起作用; 后者即系统Ⅱ 在外部K+浓度较高时起作用, 其临界点是0.2 mmol· L-1, 大约84%与78%的低亲和性及高亲和K+的吸收由AKT1与AtHAK5介导, 系统Ⅰ 与Ⅱ 约20%的是未鉴定的组分; AtHAK5与AKT1分别主要介导高亲和性及低亲和性K+的吸收[45]。然而atakt1突变体在含N
Ahmad等[48]发现, 5%及10%PEG(polyethylene glycol)处理下, 与野生型植株(WT)相比, 超表达OsAKT1的水稻OX株系地上部及根中K+浓度及生长速率明显增加; 且10%PEG明显提高了OX株系的气孔导度, 同时OX株系体内的含水量显著高于AKT1缺失体osakt1; 控水的干旱处理下, OX株系的K+含量及生长速率明显高于Osakt1及WT株, 可见, OsAKT1可通过提高K+含量而增强植株的抗旱及渗透胁迫能力。酵母菌株G19由于介导Na+外排的ATP酶ENA1~ENA4的缺失使其对Na+具有较高的敏感性[49]。而已有研究表明, 拟南芥AtHKT1; 1被该酵母表达后, 可通过介导细胞Na+的吸收从而提高G19的Na+敏感性, 抑制NaCl条件下G19的生长[50]。而K+(1 mmol· L-1)添加不同浓度Na+(0、10、30及50 mmol· L-1)的AP(arginine-phosphate)培养基中, 与转化空载体的对照相比, 转化AtHKT1; 1的G19生长显著受抑, 而转化ZxAKT1和AtAKT1的G19长势均显著好于空载体对照, 推测在NaCl处理下, 转化霸王ZxAKT1和拟南芥AtAKT1的G19的K+吸收能力显著增强, 使得酵母细胞中积累较多的K+以显著缓解Na+对细胞的毒害, 从而提高耐盐性[30]。与此相似, 在缺失高亲和性K+吸收体系的酵母菌株10A(trk1, ura3)中也表现为耐盐性提高[51]。
盐处理下, atakt1-2株系与WT的K+吸收速率没有显著差异, 因此AtAKT1对K+积累不起作用[52]。NaCl处理可能引起质膜去极化而阻碍通道介导K+吸收[53], 实际上, 低K+加盐处理下, atakt1-2株系中的外排速率要低于野生及athak5-3株系, 因此AtAKT1可能是一个K+外排的途径。因为在低K+下, 质膜可能被超级化, 从而使通道打开。K+饥饿附加NaCl处理降低了atakt1-2株系K+外排, 使根和地上部K+浓度比其它株系的高; 无盐条件下没有发现同样的现象(质膜超极化, 高的内部K+, 低的外部K+), 可能是因为没有NaCl的情况下, 膜势为负值而避免了K+外排[52]。值得注意的是, 在这些条件下, AtAKT1的调节亚基AtKC1, 不能阻止K+通过AtAKT1的外排, 而在非极端环境条件下[54, 55], 仅缺失AtAKT1降低了根中K+的外排[52]。Nieves-Cordones[52]指出, 10 μ mol· L-1 K+下, 盐的出现(30 mmol· L-1 NaCl)降低了AtHAK5的表达并引起K+通过AtAKT1外排, 可能是因为Na+引起膜势去极化, 因此K+的净吸收受到抑制。
早期学者认为, K+通道在盐胁迫下可以介导低亲和性Na+的吸收[47, 56, 57, 58], 但首先介导低亲和性K+的吸收[21, 59, 60]。Amtmann和Sanders[61]研究表明, 在低Na+浓度条件下没有明显的Na+流通过内整流K+通道。然而, 这些通道在高盐条件下可介导相对量Na+和K+的吸收[61]。TEA+(tetraethylammonium chloride)被认为K+通道的专一性抑制剂[32, 62, 63]。在水稻中发现, 耐盐性品种对Cs+和TEA+(K+通道抑制剂)并不敏感, 而Cs+和TEA+的存在条件下, 盐敏感品种细胞质中Na+浓度则均显著降低[64]。对2个不同耐盐性的水稻品种进行比较分析[24]发现, 150 mmol· L-1 NaCl处理48 h后, IR29中Na+浓度约为1 400 μ mol· g-1, 而K+浓度与对照相比增加了50%; Pokkali中Na+浓度约为200 μ mol· g-1, K+浓度则约为对照的50%; 有趣的是, 盐处理下OsAKT1在盐敏感品种IR29中表达丰度显著高于耐盐品种Pokkali, 由此表明, OsAKT1在盐敏感型品种中可能参与Na+吸收。此外, Voigt等[65]认为TEA+和Cs+能够抑制豇豆(Vigna sinensis)体内Na+的积累, 这表明K+通道在豇豆低亲和性Na+吸收方面起作用。Wang等[66]研究发现, 高盐(150 mmol· L-1 NaCl)下, 抑制剂Cs+和TEA+能够显著抑制盐生植物盐地碱蓬根部的22Na+内流, 由此推测AKT1可能具有介导高盐条件下盐地碱蓬Na+的吸收的功能。Mori等[67]对碱蓬研究发现, 培养基中含有5 mmol· L-1 K+时, Na+的吸收会显著降低, K+浓度高于5 mmol· L-1时, Na+的吸收维持不变, 当K+浓度为50 mmol· L-1时, 表现为小幅度降低, 因此认为, 碱蓬中至少存在两种Na+的吸收途径, 即对K+敏感及不敏感的两条途径。其中途径2介导对外界K+不敏感的Na+的吸收途径, 且20及100 mmol· L-1 NaCl处理下, TEA+的添加显著降低了各组织中的Na+浓度, 因此推测其可能由K+通道介导。小麦根中的TaAKT1 mRNA水平在缺K+的条件下上调, 与此同时, K+饥饿引起瞬间的Na+电流, 说明TaAKT1在K+饥饿条件下介导Na+吸收[26]。然而, 拟南芥中提高细胞质中Na+的浓度抑制了AKT1对K+的吸收[68]; 在中度盐胁迫(50 mmol· L-1 NaCl)下, 拟南芥atakt1-2突变体[16]与野生型植株相比, 其Na+浓度及净吸收速率均没有显著性差异[52], 且一些AKT1通道的基因(MKT1, OsAKT1及AKT1)表达在盐胁迫下会下调[42, 69, 70]。AKT1类通道由于对K+具有高的特异选择性不可能介导其它阳离子如Na+的吸收[71]。Ardie等[27]研究表明, 小花碱茅PutAKT1在拟南芥中超表达后, 盐胁迫下转基因植株的Na+含量下降的同时K+含量却明显提高。由此可见, 内整流K+通道是否参与Na+的吸收仍存在争议。
外界K+浓度降低时, 内整流K+通道可能具有K+传感的作用[72]。拟南芥根部的AKT1可能为K+传感器[73]。首先, AKT1是主要的内整流K+通道, 优先在拟南芥的根部表达且定位在表皮细胞的质膜上[18, 21]。其次, AKT1介导K+的吸收, 表现为典型的双亲和性特性[18, 20]。AKT1能感知[K+]ext的波动并以依赖[K+]ext的方式转变其吸收动力学位点, 位点的转变与AKT1的磷酸化水平有关, 而磷酸化水平又由CIPK23调控[38, 74]。同样, N
此外, 还有一个证据支持内整流K+通道AKT1具有K+传感器作用。有研究表明, 植物应对K+亏缺的反应是皮层细胞的膜超极化, 但是在akt1的突变体中这种超极化会完全消失[18, 20]。进一步通过异源表达发现, 当[K+]ext降低至微摩尔浓度时卵母细胞表达AKT1通道会显著地转变为超极化[13]。这些结果说明, AKT1可感应[K+]ext的变化进而影响定位在质膜上由H+-ATPases调控的膜电势[75, 76]。可见在AKT1与H+-ATPases间可能存在着一个通讯机制。近期的研究表明, K+可能结合H+-ATPases的C-末端并作为H+-ATPases解偶联ATP水解活性和质子泵活性一种内在的负调节器[77]。进一步的假设认为K+营养亏缺影响了AKT1介导的K+吸收而引起[K+]cyt的减少, 进而质膜附近减少的[K+]cyt会激活H+-ATPases的活性, 因而引起质膜超极化而介导的K+吸收的K+通道或载体被激活[73]。
近期的研究表明, CIPK23-CBL1/9复合物可激活根中AKT1从而增加K+吸收[38]。此外, CIPK-CBL复合物具有调控气孔运动及植物蒸腾作用[78]。AKT1的突变使植物对水分胁迫的响应增强, 水培实验表明, 当添加PEG时水势降低, akt1成株与野生型相比其水分损失较少, 并表现为蒸腾作用降低, 水分消耗减小, 最终, 在响应脱落酸(ABA)时其akt1植株气孔迅速关闭; 在cipk23植株中也发现类似的现象[79]。cipk23和akt1植株对水分胁迫反应的相似性表明受CIPK23调控的AKT1也可能参与气孔关闭, 并且在水分胁迫下对植株的生长具有抑制作用[79]。
K+是植物生长所需的大量营养元素, 约占植物干重的10%[71, 80]。K+是构成渗透势的重要成分, 在维系细胞正常活动所必需的跨膜电位中有不可替代的作用, 并作为多种酶的激活剂参与植株体内重要代谢[81]。内整流K+通道AKT1具有介导K+吸收、K+传感器及参与调控气孔运动及植物蒸腾作用的功能, 此外, 盐胁迫条件下AKT1还可能参与Na+吸收。因此, 基于目前的研究现状, 今后对AKT1的研究可从以下几个方面展开:1)一种生理现象的出现, 往往是多种蛋白互作的结果, 选取典型植物, 采用蛋白组学等方法深入研究K+、Na+吸收及转运的协同调控机制。2)采用基因工程技术, 将已发掘的具有同时可介导低及高亲和性K+吸收、可明显提高植株耐逆性的AKT1编码基因转入经济作物、粮食作物及饲料作物中, 提高作物对土壤中K+的利用效率及作物的产量、品质及抗性。3)在具有代表性的植物中克隆AKT1编码基因, 同时结合RNA干扰等方法揭示其在不同类型植物中的生理代谢、K+及Na+吸收及转运调控、抗逆性中的功能。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
植物内整流K+通道AKT1的研究进展
胡静, 胡小柯
Study advances of plant inward rectifying K+ channel AKT1
Hu Jing, Hu Xiao-ke