{kind=link}
{kind=link}
{kind=link}
{kind=link}
甘肃马鹿冬季牧场牧草再生性对刈割的短期响应
[马周文1  , 王宏
, 王宏2 , 于应文1 , 常生华1 , 周青平3 , 侯扶江1 ]
, 王宏]
|
|


第一作者:马周文(1991-),男,甘肃陇西人,在读硕士生,研究方向为草地-家畜互作。E-mail:[email protected]
在肃南裕固族自治县甘肃马鹿( Cervus elaphus kansuensis)不同放牧率(1.00、1.45、2.45、3.45、4.85、6.90 AUM·hm-2)的冬季牧场,研究草原生产力与群落结构对牧草最佳生长期刈割的短期响应。结果表明,1)随放牧强度增加,刈割群落物种丰富度呈先增加后减少趋势,其中4.85 AUM·hm-2放牧率下的群落物种丰富度最高;刈割显著增加草地地上生物量。2)刈割群落的牧草株高和地上生物量均表现为超补偿生长,补偿指数分别以3.45、6.90 AUM·hm-2放牧样地最高;各优势种变化趋势不同。3)刈割群落的Simpson优势度指数、Pielou均匀度指数在放牧较重样地最高;随放牧强度增加,群落Shannon-Wiener多样性指数呈先增加后减少趋势,以4.85 AUM·hm-2群落最高;且除4.85 AUM·hm-2群落外,刈割后的群落Shannon-Wiener多样性指数显著高于刈割前的( P<0.05)。4)各功能群植物对不同放牧强度响应不同。因此,适宜放牧强度下,刈割能发挥草地最大生产潜力并维持群落结构稳定性。
The study investigated the short-term effect of mowing winter pastures of deer ( Cervus elaphus kansuens) on pasture productivity and community characteristics during the best growth period, under different stocking rates (1.00, 1.45, 2.45, 3.45, 4.85, and 6.90 AUM·hm-2). We found that: 1) in mowed pastures, species richness increased with increasing stocking rate up to 4.85 AUM·hm-2 and mowing significantly increased the above-ground biomass. 2) In the mowed pastures, compensation index values indicated greater plant height and above-ground biomass. Highest compensation index was observed in the pastures grazed at 3.45 and 6.90 AUM·hm-2, respectively. The effects of stocking rates on compensation index varied with differences in the dominant plant species. 3) Simpson’s dominance and Pielou’s evenness indices for mowed pastures were generally higher in heavily grazed pastures. The Shannon-Wiener diversity index of mowed pastures increased with increasing stocking rates up to 4.85 AUM·hm-2, and the difference owing to mowing of pastures at all stocking rates, except at 4.85 AUM·hm-2, were significant. 4) We observed variation in responses among plant functional groups to pasture mowing under different stocking rates. Therefore, we suggest that stocking rates should be considered when winter pastures are mowed for maximizing production potential and maintaining the stability of community structure.
刈割是草原最重要的利用与管理方式之一。适度刈割可提高禾本科、莎草科等优良牧草分蘖、再生速率和比例, 刺激牧草补偿性生长, 从而提高草原生产力和牧草品质, 改善适口性, 促进家畜选择性采食[1, 2, 3], 优化放牧系统物质与能量分配格局, 维持群落物种多样性及结构稳定[4, 5]。刈割还可抑制毒害草生长, 改变冠层和土壤微环境, 调控温室气体排放, 增加草原生态服务功能[6, 7, 8, 9]。然而, 在放牧生态系统中, 牧草产量的季节性波动远高于畜群需求, 在牧草产量高、营养品质佳的时期, 刈割是解决季节畜牧业草畜系统相悖、加强草畜系统耦合的重要实践[10, 11]。因此, 放牧与刈割轮替、制备干草是放牧生态系统可持续的重要途径[12, 13]。
以往研究主要集中在刈割对草地生产力、土壤理化性质和群落特征等的影响[14, 15]。研究表明, 适宜的刈割时期可增加羊草(Leymus chinesis)草甸草原牧草营养物质积累[16], 高频次和高强度刈割则带走大量营养物质, 减少养分返还, 导致土壤养分循环失衡, 草原退化和生产力下降[8]。内蒙古羊草草甸群落Simpson优势度指数、Shannon-Wiener 多样性指数、Pielou均匀度指数随刈割强度的降低而减小[17]。山地高草草甸放牧-刈割草地相对单独刈割增加了土壤速效氮含量, 但低于单独放牧草地, 而其土壤有机质含量反之; 同时放牧-刈割草地的草产量是单独放牧地的2.6倍[18, 19]。放牧+长期刈割和连续放牧均增加黑麦草(Lolium perenne)+白三叶(Trifolium repens)草地植被分蘖能力, 提高群落稳定性[20]。天然草原虽然以放牧为主, 刈割为辅, 然而以往牧-刈管理制度很少在放牧强度梯度上加以研究, 至今尚未见高寒草原相关报道。
冬季牧场在夏季牧草生长峰期进行刈割, 既可以贮备干草用于放牧家畜冬季补饲, 又能保证牧草利用适宜的水热条件再生长, 提供家畜冬季放牧。但是, 高寒草原在同一生长季内刈牧兼用是否具有可行性, 对群落结构与功能有哪些作用, 尚不清楚。为此, 本研究在祁连山高寒草原甘肃马鹿(Cervus elaphus kansuensis)冬季牧场, 分析放牧强度梯度上刈割对草原生产力与群落结构的短期作用, 以期为草原合理利用提供科学依据。
研究区位于甘肃省河西走廊南部、祁连山北麓中段的肃南裕固族自治县甘肃马鹿养殖场(38.8° N, 99.6° E), 海拔2 850 m, 年均温3.6 ℃, 年均降水量260 mm, 属高寒山地半干旱气候区。降水主要集中在6月-9月, 年均蒸发量1 785 mm, 日照时数3 085 h, 无霜期127 d左右。土壤为山地栗钙土。植被一般于5月上旬返青, 6月下旬逐渐进入生长旺盛期, 9月中上旬开始枯黄。草地类型属寒温微干山地草原类, 优势物种有紫花针茅(Stipa purpurea)、赖草(Leymus secalinus )、早熟禾(Poa annua)、苔草(Carex ateriima)等, 退化区域主要有醉马草(Achnatherum inebrians)。
冬季牧场属严重退化草地, 自1998年开始, 甘肃马鹿于每年11月下旬-翌年2月放牧。通过实地调查和询问牧民, 确认马鹿放牧采食路线, 样地以牧场进出口为起点, 向外呈放射状地形成放牧率由高到低的牧压梯度:6.90、4.85、3.45、2.45、1.45和 1.00 AUM· hm-2。放牧率(AUM)以单位放牧地面积上家畜单位与放牧时间(月, month, M)的乘积表示, 其中1头成年马鹿为1个家畜单位(animal unit, AU), 各年龄阶段马鹿根据体质量换算为家畜单位[21, 22], 分别对应在距牧场出入口0、300、600、900、1 200和1 500 m处设置的6个样地, 每样地面积约1 hm2。
采用限定随机取样法[23], 于1999年7月下旬在每个放牧梯度样地刈割4个1 m2样方, 留茬高度为2 cm左右, 1个月后调查刈割与未刈割群落的植被生长特征(株高、密度、盖度、地上生物量)。
1.3.1 补偿指数的计算 放牧群落补偿生长模式用补偿指数G/C表示[24]。G为8月底刈割处理样地的株高、密度或盖度的补偿量, C为对照样地8月底的株高、密度或盖度。若G/C> 1, 为超补偿生长; 若G/C=1, 为等补偿生长; 若G/C< 1, 为低补偿生长。
1.3.2 草地群落物种多样性(α 多样性指数)的计算 根据测定各植物种的密度、高度、盖度, 分别计算群落α 多样性指数, 计算公式如下[25, 26]。
重要值(IV)= (相对盖度+相对高度+相对密度)/3;
Shannon-Weiner 指数:H'=-∑ Pilg(Pi);
Simpson 指数: D=1-∑
Pielou均匀度指数:E=H'/lnS.
式中:S为每个样方中的物种数目; Pi=IV/∑ IV。
采用SPSS 17.0软件对群落物种丰富度、地上生物量、补偿指数、α 多样性指数、功能群重要值进行了方差分析(LSD), 用Microsoft Excel软件进行图表绘制。
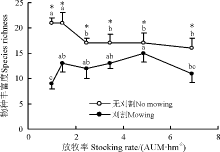
随放牧强度增加, 刈割群落的物种丰富度整体呈上升趋势, 4.85 AUM· hm-2群落显著高于6.90和1.00 AUM· hm-2群落(P< 0.05)。无刈割群落物种丰富度则随放牧强度增加一直递减, 重度放牧区显著低于轻度放牧区(P< 0.05)。除4.85 AUM· hm-2群落外, 群落物种丰富度对刈割较敏感(图1)。
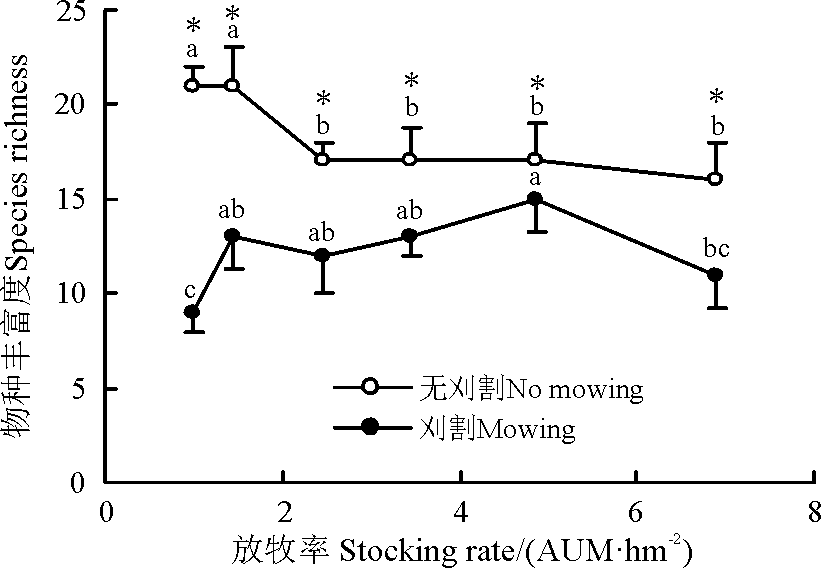 | 图1 刈割对不同放牧强度下草地物种丰富度的影响* 表示同一放牧率刈割与无刈割间差异显著(P< 0.05); 不同小写字母表示同一刈割(无刈割)处理不同放牧率间差异显著(P< 0.05)。下同。Fig. 1 Effect of mowing on species richness of grassland under different stocking rates* the same stocking rate at indicates significant difference between mowing and no mowing pastures at the 0.05 level; different lowercase letters indicate significant differences among different stocking rate under mowing or no mowing condition at the 0.05 level. The same as below. |
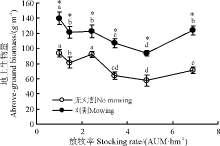
随放牧强度增加, 刈割草地地上生物量表现为先减少后增加趋势, 1.00 AUM· hm-2群落最高, 4.85 AUM· hm-2群落最低, 无刈割(除2.45 AUM· hm-2)与刈割群落变化趋势基本相同且二者差异显著(P< 0.05)(图2)。
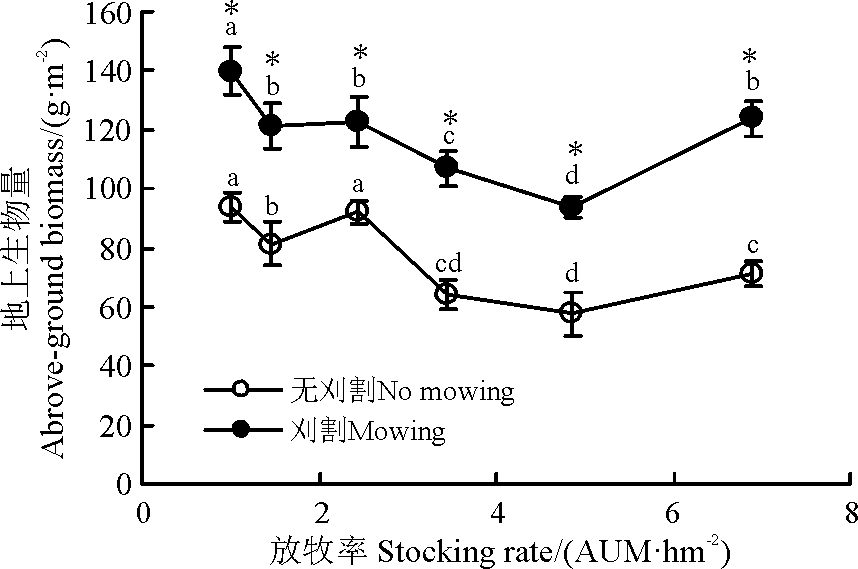 | 图2 刈割对不同放牧强度下草地地上生物量的影响Fig. 2 Effect of mowing on above-ground biomass of grassland under different stocking rates |
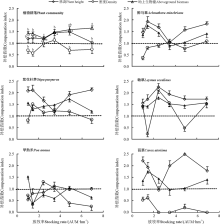
不同放牧强度下, 植物群落及其优势种补偿指数具有明显差异(图3)。刈割群落株高均表现为超补偿生长, 4.85、3.45 AUM· hm-2样地显著高于6.90和2.45 AUM· hm-2样地(P< 0.05)。除3.45 AUM· hm-2群落外, 群落密度补偿指数显示群落密度为低补偿生长, 且1.45 AUM· hm-2群落最小, 显著低于3.45 AUM· hm-2群落(P< 0.05)。群落地上生物量补偿指数显示群落地上生物量为超补偿生长, 随放牧强度增加表现出先减少后增加趋势, 放牧较重的6.90 AUM· hm-2群落显著高于其它群落(P< 0.05)。刈割不利于早熟禾的短期生长, 而促进紫花针茅的再生长, 不同放牧强度下的响应不同; 适度放牧强度下刈割可抑制醉马草的再生速率, 但利于赖草的再生长。
 | 图3 植物群落及其优势种补偿指数对放牧强度的响应Fig. 3 Response of plant community and its dominant species compensation index to stocking rates |
高寒草原物种多样性对刈割及放牧强度具有明显的响应(表1)。无刈割草地群落, Simpson优势度指数、Shannon-Wiener多样性指数、Pielou均匀度指数在不同放牧强度间差异不显著(P> 0.05)。刈割群落, Simpson优势度指数在3.45和2.45 AUM· hm-2群落较高, 不同放牧强度间差异不显著(P> 0.05); Shannon-Wiener 多样性指数随放牧强度增加呈先增加后减少趋势, 4.85 AUM· hm-2群落显著高于其它群落(除3.45 AUM· hm-2); 且4.85 AUM· hm-2群落最高, 显著高于1.00 AUM· hm-2群落37.06%(P< 0.05); Pielou均匀度指数在4.85 AUM· hm-2群落最高, 1.45 AUM· hm-2群落最低, 但不同放牧强度间差异不显著(P> 0.05)。刈割群落的Simpson优势度指数、Shannon-Wiener 多样性指数、Pielou均匀度指数均较无刈割减小, 且只有Shannon-Wiener 多样性指数在刈割与不刈割间差异显著, 除4.85 AUM· hm-2样地外, 刈割样地均显著低于未刈割(P< 0.05)。Simpson优势度指数以4.85、1.00 AUM· hm-2群落最敏感, 相对无刈割分别减少了13.64%和13.33%; Shannon-Wiener 多样性指数以1.00 AUM· hm-2群落最敏感, 相对无刈割减少了32.81%; Pielou均匀度指数以1.45 AUM· hm-2群落最敏感, 相对无刈割减少了8.75%。草地群落多样性较重度放牧样地较高。
| 表1 刈割对不同放牧强度下草地群落α 多样性指数的影响 Table 1 Effect of mowing on α diversity index of grassland community under different stocking rates |
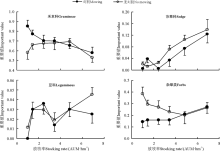
各植物功能群对放牧强度的响应不同(图4)。刈割后随放牧增强, 禾本科重要值呈逐渐减小的趋势, 相比无刈割轻度放牧时其功能群重要值提高; 杂类草、莎草科功能群重要值增加, 相比无刈割处理, 轻度放牧区的功能群重要值降低; 豆科功能群重要值基本无变化。不同放牧强度下, 禾本科功能群重要值高于其它功能群; 但重度放牧区, 禾本科功能群的重要值降低, 而其它功能群的重要值增加。
 | 图4 刈割对不同放牧强度植物群落功能群的影响Fig. 4 Effect of mowing on functional groups of plant community with different stocking rates |
牧场刈牧兼用模式有助于改善草原的健康状况[27]。不同的刈牧方式、时间和制度等影响草地群落的结构和功能, 进而影响草地的经济效益及生态服务功能[28]。不合理的利用方式致使草地生产力下降, 毒杂草等的重要值增加, 饲草营养品质降低, 影响家畜采食, 进而使群落结构恶性演替, 最终导致草地退化[20, 29, 30]。
刈割群落的物种丰富度短期内在重度和轻度放牧群落减少, 与无刈割群落相比, 前者地上生物量显著增加(图1、图2)。因为适度刈割群落的优势种的盖度降低, 促进杂类草繁殖和生长, 导致群落物种丰富度和地上生物量增加[31]。也有研究显示, 中度及适度刈割频次能提高再生速率进而提高草产量[32]; 刈割可减少群落物种数, 主要是由于刈割时间过晚影响物种对营养物质积累, 进而影响物种的越冬[33]或刈割减少了土壤种子库[19]。
不同科属植物补偿机理不同, 导致不同放牧强度群落产生不同的补偿效应[34]。适宜放牧率条件下, 群落在刈割后有利于草原群落株高、密度和地上生物量短期的超补偿生长, 从而使草地生产力显著提高(图3), 其可能原因是适宜放牧率下, 不同科属植物的组成比例、空间构建格局比较合理, 在刈割后其生长速率在短期容易得到补偿[35]。由于重度放牧群落中菊科[紫菀(Aster tataricus)等]和毒杂草类植物[醉马草、银灰旋花(Convolvulus ammannii)等]较多, 所以其株高超补偿指数较低, 密度为低补偿生长, 而地上生物量表现为较高超补偿生长; 轻度放牧群落由于禾本科牧草[紫花针茅、赖草、冰草(Agropyron cristatum)等]比例大, 所以其株高和地上生物量超补偿指数较低, 密度表现为低补偿生长。而付秀琴和王梅[34]、樊江文等[36]研究认为, 草地补偿模式是由根系生长的独立性与牧草耐牧性(耐干扰性)决定。
短期刈割影响下, 草地群落的物种多样性Shannon-Wiener多样性指数受影响较大, 且Simpson优势度指数、Shannon-Wiener 多样性指数、Pielou均匀度指数均在相对较重度群落最高(表1)。说明刈割后草地植被物质和能量的空间利用格局得到重新分配, 影响群落的物种多样性[4, 5]。但对于灌草丛草地刈割可促进土壤种子库的种子萌发, 增加物种多样性[37]。另外, 草地生态系统功能依赖于生态系统中物种功能群属性及其功能性状的组成[27]。刈割后禾本科短期表现为重度放牧抑制型功能群, 杂类草、莎草科表现为重度放牧促进型功能群, 豆科表现为轻度、重度放牧促进型功能群(图4), 这与降低放牧强度有利于提高青藏高原高寒嵩草草甸中禾本科和莎草科在群落中的地位[38]结果一致。其原因是不同处理下各功能群反馈速率和累积效应不同所致[27, 34, 39]。
综上所述, 同一生长季内实现刈牧兼用, 较轻放牧群落较高放牧率群落表现出较大的可实施性。可见, 合理的“ 放牧-刈割制度” 能提高草地生产力, 有利于物种丰富度的维持、物种多样性的提高及功能群结构的优化, 并对生态系统稳定维持具有重要作用。本研究仅为“ 放牧-刈割” 制度在一个生长季节的短期响应, 其在刈割高度和刈割强度的长期效应还有待进一步研究。
(责任编辑 武艳培)
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|