
{kind=link}
山西浊漳河北源湿地草本植物群落优势种生态位与种间关系
[王昱1  , 毕润成
, 毕润成1 , 茹文明2, 3 ]
, 毕润成]
|
|


第一作者:王昱(1992-),女,山西太原人,在读硕士生,主要从事植物生态学研究。E-mail:[email protected]
以山西浊漳河北源湿地草本植物群落调查为基础,通过生态位指数测度、方差比率法、χ2检验、联结系数AC、Pearson相关系数检验对16个优势种群进行了生态位和种间关联的分析。结果表明,1)藨草( Scirpus triqueter)、头穗莎草 ( Cyperus glomeratus)、水蓼( Polygonum hydropiper)的生态位宽度值相对较大,在研究区域内具有泛化种倾向;2)120个种对间的生态位重叠指数低,55%的种对之间不存在重叠,种间竞争弱;3)群落内植物整体关联度为显著负相关( P<0.05),χ2检验与Pearson相关系数检验的正负关联比(分别为0.52和0.15)均小于1,负关联的种对数多于正关联,且大多数种间关联程度不显著( P>0.05),说明浊漳河北源湿地草本植物群落结构简单,物种间相互作用较弱,处于演替初期阶段;4)物种间的生态位重叠指数与联结系数AC、Pearson相关系数存在着显著的正相关关系( P<0.05),且Pearson相关系数的拟合效果优于AC联结系数。本研究有利于更好地认识优势种种间的相互关系及群落的动态特征,了解浊漳河北源湿地利用现状,为湿地生态恢复提供理论依据。
Based on sample investigations of wetland herb communities in the northern branch of the Zhuozhang River Watershed, we analyzed interspecies relationships and niche characteristics for 16 main species. Important findings were as follows: 1) Scirpus triqueter, Polygonum hydropiper, and Cyperus glomeratus showed tendencies of generalized species such as the largest niche breadth among dominant species; 2) a niche overlap of 55% was observed in 0 of 120 species pairs, and lower niche overlap between species indicated weaker interspecific competition; 3) Overall association of phytocoenosium was significantly negative correlation at the 0.05 level, the ratios of positive and negative associations in χ2-tests and Pearson correlation coefficients were 0.52 and 0.15; therefore, the negative associations were more than the positive associations, and most interspecific associations were not significant, suggesting that wetland herb communities structure is simple, the interaction between species is weak, the communities are still under dynamic succession; 4) both association coefficients (AC)and Pearson correlation coefficients indicated niche overlap, with the fitting effect of Pearson correlation coefficient showing a stronger positive association than the AC. To provide a theoretical foundation for ecological restoration in the norther branch of the Zhuozhang River Watershed, this study integrated niche characteristics and interspecific associations to understand the dominant species, their interrelations, dynamic characteristics of the communities and current wetland utilization.
生态位可以描述物种在群落或生境中的地位和作用, 反映物种对环境资源的需求[1, 2]。分析物种的生态位特征有利于认识自然群落中物种之间的竞争与共存关系[3], 进而揭示群落结构和功能、推测群落的演替进程。种间关系是不同物种之间的相互关联性在空间分布上的表现[4], 能够反映种间的相互作用及物种与环境间的耦合关系[5], 反映物种在不同生境中相互影响相互作用形成的有机联系及物种间相互吸引或相互排斥的性质[6, 7]。通过研究物种生态位与物种间的相互关系, 了解各物种在群落中的功能地位、相互关系, 对植被的利用与保护具有重要意义。湿地被称为“ 地球之肾” 、“ 淡水之源” 、“ 天然物种库” .它作为世界上最重要的生态系统之一, 在调节气候、维持区域生态平衡和保护生物多样性等诸多方面有着重要作用[8, 9]。湿地植被是湿地生态系统的重要组分[10], 在湿地生态系统中发挥着关键作用, 有着补给水源、调节湿地小气候、指示湿地环境变化的功能。浊漳河湿地属内陆半干旱湿地, 长期以来在饮用水源、农业用水、采煤发电等方面发挥着较大的经济作用, 同时也对改善当地环境质量、稳定区域气候做出了积极的生态贡献, 是山西省稀缺的湿地资源。然而, 受流域内经济发展和人口激增的影响, 浊漳河流域人为污染加剧, 河流水质恶化、湿地面积萎缩、湿地植被退化, 湿地生态环境承载力不断下降, 引起了相关学者的广泛关注。但是对浊漳河流域的研究多集中在水资源利用[11]、地表水环境[12]与水污染方面[13], 缺少对湿地植被的研究。近两年, 王烨等[14]和金山等[15]对浊漳河干流、南源植被开展了部分研究, 对浊漳河流域进行了群落类型的划分、物种多样性和种间关系及功能群的分析。本研究在上述研究的基础上, 探讨浊漳河北源湿地草本植物群落优势种间相互依存、相互制约的生态关系和植物对生态环境的适应能力, 以期为浊漳河北源湿地群落的恢复与保护措施提供依据, 对内陆湿地生态的开发与维持、利用与恢复具有一定的理论意义。
浊漳河北源是浊漳河三源之一, 发源于晋中市榆社县柳树沟, 总流域面积3 791.64 km2, 主河道长116 km。北源干流自晋中市榆社县流入长治市武乡县, 过关河水库后, 流向由向南改为向东南, 流至襄垣县小嶕村南汇入浊漳河干流。研究区地跨晋中、长治两市, 覆盖榆社、武乡、襄垣三县, 地理范围为36° 38'72″-37° 14'28.2″ N, 112° 55'13.4″-113° 36'72″ E。总体地势西北高, 东南低, 平均海拔979.6 m, 属于温带大陆性季风气候区, 年均气温8 ℃左右, 年均降水量550~580 mm。流域内大部分地区经济欠发达, 土地多被开垦为农田, 森林面积少, 植被覆盖率低, 属黄土丘陵区, 黄土结构松散, 下渗强度大, 易被冲蚀, 水土流失严重[11]。主要草本植物:芦苇(Phragmites communis)、小香蒲(Typha minima)、泽泻(Alisma orietalecsam)、藨草(Scirpus triqueter)、水莎草(Juncellus serotinus)、球穗莎草(Cyperus difformis)、异型莎草(Cyperus difformis)、稗(Echinochloa colonum)、水蓼(Polygonum hydropiper)、酸模叶蓼(Polygonum lapathifolium)等[16]。
2016年7-8月, 在浊漳河流域湿地植物生长的茂盛时期, 以湿地草本植物为研究对象, 根据浊漳河流域的植物群落类型、湿地类型(河流湿地、水库湿地、沼泽湿地、河漫滩草甸及河岸湿草甸)以及人为干扰的程度, 在距离北源河道0-4 m内选取能客观反映浊漳河湿地植物现状的样地进行调查。采样范围包括被调查湿地内有沉水植物生长的水体、水陆过渡地带以及长期或季节性受湿地水分影响的区域。样地布设沿浊漳河北源河道走向选取, 样地概况如表1所列。在样地的4个角(或四角及中心)设置4~5个1 m× 1 m的草本样方, 共得到20个样地, 81个样方。
| 表1 样地概况 Table 1 Sample plot |
记录样方内植物的种名、高度和盖度, 生境因子记录经纬度、海拔、坡度、坡向等。
1.3.1 重要值(IV)
IV=(相对高度+相对频度+相对盖度)/3。
在所调查的81个样方中共记录物种48个, 根据各个物种重要值的大小及综合各物种出现的频度(剔除频度< 5%, 频度以物种出现的样方数目占样方总数的百分比来测算), 选取出16个优势种, 进行生态位与种间关联的指标计算。
1.3.2 生态位相关指数测度
1)生态位宽度
生态位宽度的计算以Shannon-Wiener指数测定[17]。计算公式如下:
Bi=-
式中:Bi表示种i在r个资源下的生态位宽度, Pij表示种i对第j个资源的利用占全体种群对第j个资源利用的频度。
2)生态位重叠
优势种群生态位重叠(NO)的计算公式以重叠指数Pianka[18, 19]为标准:
NO=
式中:P为物种重要值, N为样方数, Pij和Pkj分别表示在样方j中种i和种k的重要值。
1.3.3 总体相关性检验 要从整体上反映群落优势物种的关联性需要进行总体相关性的检验。检验方法是计算方差比率值(VR)[20]:
VR=
式中:S为物种总数, N为样方总数, t为样方中物种的平均数, ni为物种i出现的样方数目, Tj为样方j内出现的物种数目。
若VR=1, 种间整体无关联; VR> 1, 种间关系在整体上呈正相关关系; 若VR< 1, 种间关系在总体呈负相关关系。W是检验VR显著性的指数, 当W<
1.3.4 χ 2检验 将Yates的连续性校正公式[21]用于检验χ 2 :
χ 2=
式中:两个物种都出现的样方数、只出现一个物种的样方数、两物种都未出现的样方数分别用a、b、c、d进行表示, N为样方总数。以ad与bc乘积的差值(ad-bc)判断正负关联, 正关联差值大于0, 负关联小于0。以计算出的χ 2判断显著程度, χ 2< 3.841, 种间关联性不显著(P> 0.05); 3.841< χ 2< 6.635, 种间显著关联(P< 0.05); χ 2> 6.635, 种间呈极显著相关(P< 0.01)。
1.3.5 联结系数AC 以2× 2连列表的形式, 计算联结系数[22]:
AC=(ad-bc)/(a+b)(b+d), ad≥ bc;
AC=(ad-bc)/(a+b)(a+c), bc> ad, d≥ a;
AC=(ad-bc)/(d+b)(d+c), bc> ad, d< a。
当-1≤ AC≤ 1, AC=0时两个物种彼此独立; AC的值越趋近于1, 物种间的正联结性越强, 接近-1则负联结性越强。
1.3.6 Pearson积矩相关系数(rik)按照如下公式计算:
rik=
式中:xij和xkj为种i和种k在样方j中的秩(按重要值计算)。
1.3.7 数据分析 对优势种的生态位重叠指数值与联结系数AC、Pearson相关系数进行相关性分析。本研究使用Excel、SPSS 17.0和R进行数据处理。
在调查的81个样方中, 浊漳河北源流域共出现草本植物48种, 群落覆盖度为60%~70%。生态位宽度最大的物种为藨草, 其次是头穗莎草、水蓼; 生态位宽度最小的物种为球穗扁莎、其次是荩草(Arthraxon hispidus)、萤蔺(Scirpus juncoides)。藨草的重要值和生态位宽度均最大, 表明藨草在草本群落中占据绝对优势, 在群落构建过程中起主导作用(表2)。
| 表2 浊漳河北源优势种重要值及生态位宽度 Table 2 Importance values and niche breadths of the dominant species in the norther branch of the Zhuozhang River wetland |
120个种对中, 生态位重叠值> 0.40的仅有1对, 0.2~0.4的有4对, < 0.20的有115对, 其中等于0的有66对, 占总对数的55%。表明浊漳河北源湿地草本植物优势种之间生态位重叠程度低, 各物种对资源利用的相似性较小, 物种生物学和生态学特性存在差异, 竞争关系不显著(P> 0.05)(表3)。
| 表3 浊漳河北源草本植物群落优势种生态位重叠 Table 3 Niche overlap of the dominant species in herbaceous communities in the northern branch of the Zhuozhang River wetland |
16个优势种的总体联结性指数VR< 1, 主要种群间总体呈关联。方差检验比例系数W值在区间[
| 表4 浊漳河北源优势草本植物总体关联性检验 Table 4 Dominant plant species general associativity test for communities in the northern branch of the Zhuozhang River wetland |
2.4.1 联结系数AC 从联结系数AC来看(表5), AC值在[-1, -0.4)、[-0.4, 0.4)、(0.4, 1]的种对数分别为4、67、49个, 占比55.83%、40.83%, 3.33%。由于在AC值计算方法中, 两物种没有共同出现的样地其值必为0, 使得计算所得极显著负相关的物种对数增多。故进行χ 2检验来弥补这一算法的不足。
| 表5 联结系数范围及相应种对数 Table 5 AC values of inter-specific associations and the corresponding number of species pairs |
2.4.2 χ 2检验 根据Yates公式[21]校正后的χ 2检验结果(表6)可知, 呈极显著(P< 0.01)正相关关系的种对包括藨草(1)与香蒲(4)、藨草(1)与头穗莎草(8)、狼尾草(2)与艾(16)、稗(5)与狼杷草(12)、狐尾藻(10)与菹草(11); 呈显著(P< 0.05)正相关关系的种对包括水蓼(3)与稗(5)、香蒲(4)与芦苇(7); 旋覆花(13)与艾(16); 呈极显著负相关关系的种对包括藨草(1)与狼尾草(2)、藨草(1)与艾(16)、狼尾草(2)与头穗莎草(8)、头穗莎草(8)与艾(16); 呈显著负相关关系的种对包括藨草(1)与狐尾藻(10)、藨草(1)与菹草(11)、藨草(1)与旋覆花(13)、头穗莎草(8)与菹草(11)。
| 表6 16个优势种的χ 2校正检验半矩阵 Table 6 Semi-matrix of χ 2 test results for 16 dominant species |
在16个优势种组成的120个种对中, 正相关的种对有41个, 负相关的种对有79个, 正负关联比为0.52; 其中呈极显著(P< 0.01)相关的种对有9个, 显著(P< 0.05)相关的种对有7个, 不显著(P> 0.05)相关的种对有104个, 分别占总对数的7.5%、5.83%、86.67%。总体上看, 120个种对中, 负相关的种对数大于正相关的种对数, 这一点与多物种间的总体关联性检验所得结果一致; 另外, 显著相关的种对数远小于不显著相关的种对数, 种对之间植被盖度值相互变化趋势不明显, 空间相对分布位置关系不明确。表明多数物种间尚未产生竞争关系, 浊漳河北源湿地草本植物群落种间的联结性弱。
2.4.3 Pearson相关系数检验 Pearson相关系数检验半矩阵如表7所列, 极显著(P< 0.01)和显著(P< 0.05)正相关的种对各1个, 分别是狼尾草(2)与艾(16), 狐尾藻(10)与菹草(11); 没有极显著负相关、显著负相关的种对。120个种对中, 正相关的种对有个16个, 负相关的种对有104个, 正负关联比(0.15)远小于1。其中极显著相关的种对1个, 显著相关的种对1个, 不显著(P> 0.05)相关的种对158个, 分别占总种对数的0.62%、0.62%、98.76%。160个种对中, 负相关的种对数多于正相关的种对数, 负相关的种对数出现的频率很高, 但却没有极显著和显著相关的种对数, 表明该湿地的草本植物群落结构不够稳定, 很容易受到干扰而产生变化, 群落尚处在演替初期阶段, 且仅有2对达到显著程度, 优势种种间联结松散, 与χ 2检验的结果基本一致。
| 表7 16个优势种的的Pearson相关系数检验半矩阵 Table 7 Semi-matrix of Pearson’ s correlations among 16 dominant species |
生态位重叠与物种间的AC联结系数、Pearson相关系数之间正相关关系显著(P< 0.05)(图1)。Pearson相关系数与生态位重叠指数的回归方程为y=0.056 1+0.774x+2.200 8x2-3.686 5x3, (R2=0.877 9, P< 0.05); 联结系数AC与生态位重叠指数为y=0.051 4+0.175 6x+0.202 6x2+0.077 4x3, (R2=0.698 1, P< 0.05); Pearson相关系数与生态位重叠指数间的相关指数R2更接近于1, 拟合效果较AC联结系数更好。
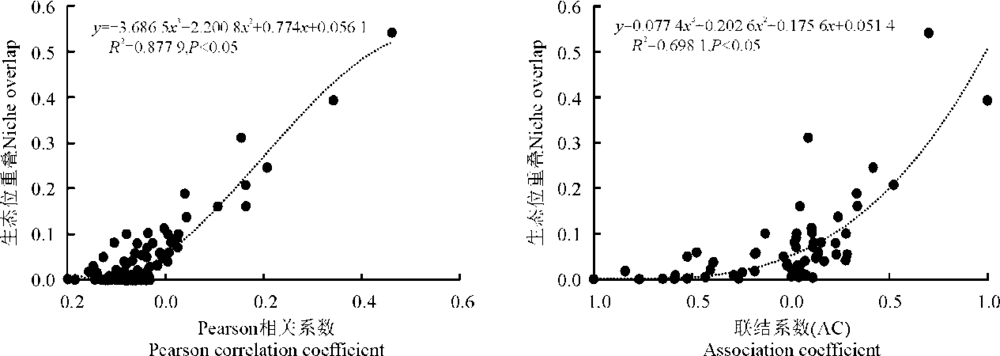 | 图1 种间关系与生态位重叠的回归分析Fig. 1 Regression analysis of interspecific associations and niche overlap |
生态位宽度可以量化物种对资源与环境的利用程度, 物种的生态位越宽, 对环境的适应能力越强, 特化程度小, 倾向于泛化种, 常是群落中的优势物种[23]。本研究中, 藨草、头穗莎草、水蓼、芦苇较宽的生态位表明了它们在研究区的泛化种倾向; 藨草的重要值和生态位宽度值最大, 表明该物种在研究区占据绝对的优势地位。
生态位重叠能够反映出种群之间的竞争、共存机制。生态位重叠指数高的种群对环境资源的生态需求相似, 容易因为资源的占用导致竞争, 而竞争又是群落演替的动因之一[24]。研究表明, 生态位宽度都较大或生态位宽度较窄却出现在少数同一资源位上的两个物种生态位重叠较高[25, 26]。在调查的120个种对中, 有66对的生态位重叠指数为0, 占总数的55%, 说明浊漳河北源湿地草本植物群落种间生态位重叠指数低, 竞争关系弱; 群落中, 狼尾草和艾的生态位宽度和生态位重叠大, 二者对环境的适应能力及资源的利用能力强, 易在环境中出现、生存; 狐尾藻-菹草、水蓼-旋覆花、香蒲-芦苇的生态位宽度窄但生态位重叠值大, 这是由于物种对环境和资源的需求相似性高, 即使对资源和环境条件有所要求, 但环境条件满足会同时出现。
物种的生态学特性、生态适应性和生态位分化使得种间具有了不同的正、负关系。这些关系是植物对环境综合生态因子反映的差异[27]。正关联的种对对环境资源的需求相似, 而负关联的种对则利用不同的空间资源或在竞争中互相排斥[28]。研究区草本植物群落优势种总体上呈负关联, 物种间总体相似性较低, 这与物种间生态位重叠指数偏低相吻合。一般来说, 物种间会随着群落的发展演替形成稳定共存的关系而趋向于正相关。本研究中的χ 2检验与Pearson相关系数检验结果表明, 120个种对的正负关联比为0.52和0.15, 均小于1, 且大多数种对间呈不显著相关。这说明优势种间相对独立, 群落易受到外界干扰而发生波动[29]。因此, 可以推测浊漳河北源湿地草本植物群落种间相互作用较弱, 群落整体发展水平较低, 正处于演替的前期阶段。
对浊漳河北源湿地草本植物群落优势种的生态位重叠与联结系数AC、Pearson相关系数做回归分析得出, 生态位重叠与AC联结系数、Pearson相关系数之间表现为显著的正相关; Pearson相关系数与生态位重叠间的回归模型拟合效果更好。在回归方程中, Person相关系数在[-0.1, 0]的种对数目最多, 这些种对的生态位重叠也较低, 范围为[0, 0.1], 这表明大多数种对呈不显著的负相关, 各物种相互独立, 尚未产生竞争, 不具备物种间稳定共存的特征。相关系数在0.2以上的种对数有6对, 这些种对间的生态位重叠值范围为[0.2, 0.6], 正相关的种对生态位重叠值大, 反映出对生境要求的一致性。如藨草和球穗扁莎, 狐尾藻和菹草, 狼尾草、艾、旋覆花、假苇拂子茅、荩草。藨草和球穗莎草为莎草科湿生植物, 生态学特性相似; 狐尾藻不耐寒, 好温暖水湿的环境, 菹草为多年生沉水草本植物, 二者都属湿生植物; 狼尾草、艾喜温湿, 亦能耐旱耐阴, 旋覆花多生于草地、河岸等比较湿润的环境, 假苇拂子茅、荩草多生于山坡草地阴湿之处, 为中生草本植物, 均为湿中、中生植物。对水分、温度、光照的需求是湿地植被间生态位重叠及种对间正负关联的关键。由于群落结构不成熟, 即使对资源的利用相似性高, 尚能“ 和平共处” , 资源竞争不显著。如果群落进行正向演替, 则会有一方占据优势或通过生态位分化达到稳定共存。
本研究采用生态位特征与种间关系相结合的方法对浊漳河北源湿地草本植物群落优势种进行分析, 结果表明:大多数种群间的生态位重叠较小, 种间竞争较弱, 物种之间彼此独立。物种总体相关性表现为显著的负联结; AC联结系数、χ 2检验、Pearson相关系数检验的正负关联比均小于1, 且大多数种对表现为中性联结, 说明物种间的相互作用较弱, 群落整体发展水平较低, 群落结构相对不稳定, 容易受到外界因素的干扰。生态位重叠指数与联结系数AC、Pearson相关系数之间具有显著的正相关关系, 在本研究中, 负关联的种对数居多且各物种间的生态位重叠值小的结论也反映了这种正相关关系。综合来看, 两种方法的研究结果都表明浊漳河北源湿地草本植物群落较为脆弱, 群落结构简单, 易受外界干扰, 尚处在演替初期阶段, 有着较大的相似性, 可以互相补充。鉴于上述研究结论, 对于浊漳河北源湿地流域, 应当减少湿地人为破坏, 保证湿地群落的自然演替进程, 建立湿地自然保护区, 以期更好地保护和可持续地开发利用湿地资源。
(责任编辑 武艳培)
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|