





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
AM真菌与禾草内生真菌对黑麦草抗旱性的影响
[王晓瑜 , 郭艳娥, 冯希, 施颖, 段廷玉]
, 郭艳娥, 冯希, 施颖, 段廷玉]
, 郭艳娥, 冯希, 施颖, 段廷玉]
|
|


第一作者:王晓瑜(1996-),男,甘肃民勤人,在读本科生,研究方向为草业科学基础理论。E-mail:[email protected]
通过温室盆栽试验,从植物生长、养分吸收和抗逆相关酶等方面探究了AM真菌与禾草内生真菌互作对黑麦草( Loliumperenne)抗旱性的影响,以期为充分利用两类微生物来提高植物抗逆性提供理论依据。本研究包括单独接种AM真菌、禾草内生真菌,AM真菌+禾草内生真菌以及无AM真菌和无禾草内生真菌4种处理,黑麦草分别由含有禾草内生真菌(E+)和不含禾草内生真菌(E-)种子建植获得。并于黑麦草生长38 d后,按照土壤最大持水量的15%、30%、45%和60%设置4个水分梯度,共16种处理。结果表明,接种AM真菌与禾草内生真菌显著促进了45%和60%土壤水分条件下黑麦草的生长,促进了其对N、P的吸收,增强了黑麦草体内过氧化物酶活性,降低了丙二醛浓度,表明本研究所用共生微生物仅在较为干旱条件下提高了植物的耐旱性。
We established a pot experiment in a greenhouse to study the effects of arbuscular mycorrhiza(AM) fungus and a grass endophyte on perennial ryegrass drought tolerance, aiming to identify two kinds of symbioticmic roorganisms to improve plant drought stress tolerance. The AM treatments involved inoculation with or without arbuscular mycorrhizal fungus.Seeds of perennial ryegrass infected with a grass endophyte (E+) and without grass endophyte (E-) were used to establish plants with and without a grass endophyte, respectively. After 38 days of growth, ryegrass grown in the above microorganism treatments were exposed to four soil water treatments of 15%, 30%, 45%, and 60% of the maximum soil water holding capacity. The mycorrhizalfungus and grass endophyte mostly showed positive effectson plant growth and nutrition uptake in 45% and 60% soil water. The two types of symbiotic fungi not onlypromoted the growth of perennial ryegrass and enhanced shoot and root biomass but also promoted plant nitrogen and phosphorus uptake, enhanced peroxidase activity, and reduced malonaldehyde concentrations.Therefore, the two symbionts enhanced plant drought tolerance and reduced ( P<0.05) the damage to perennial ryegrass under semi drought stress.
水资源匮乏是限制植物生长和生态系统生产力的主要因素之一[1], 不仅影响植物生长和发育, 而且造成土地荒漠化及生态环境的恶化, 是引起农牧业减产的首要环境因素[2], 其影响超过其他环境胁迫的总和[3]。我国有超过一半面积的地区处于干旱或半干旱状态, 部分地区局部缺水状况较为严重, 这些地区年均降水量均在500 mm以下[4]。2016年春季, 由于受到季风影响, 我国华北大部、内蒙古、黄淮地区以及西北地区, 在不同时段内分别受到不同程度干旱气候的影响[5]。
利用微生物, 尤其是共生微生物提高植物抗旱性和修复生态系统是国际上研究的热点领域。丛枝菌根(arbuscular mycorrhizae, AM)真菌是广泛分布于自然生态系统中的一类土壤真菌, 能与绝大多数高等植物根系形成共生体[6]。AM真菌可形成连接根际土壤与根系表皮细胞的菌丝网[7, 8], 促进植物对土壤矿质元素[9]和水分[10]的吸收和利用, 改善植物营养状况, 促进生长发育[11, 12], 提高植物抗旱性。因此, 干旱环境中AM真菌与植物生长的关系倍受人们关注[13]。禾草内生真菌在草类家族中广泛分布, 在早熟禾亚科的冷季型草中分布最广[14]。禾草内生真菌作为目前国内外研究的热点, 相关研究主要集中在其提高作物抗病、促生和抗逆等方面[15], 禾草内生真菌促进植物抗旱性[14, 16, 17]和抗虫性[18, 19, 20]的研究也已有报道。禾草-内生真菌-家畜的关系, 集中反映了草地农业生态系统中, 植物与微生物, 植物与动物, 微生物与动物之间的错综复杂关系[20]。
黑麦草(Lolium perenne)作为多年生的优质禾本科牧草, 具有生长快、分蘖多、再生性好等特点[21], 是我国退耕还草和栽培草地建植的主要草种之一, 也是改良我国干旱地区生态环境、防风固沙的优良草种。黑麦草草质柔嫩多汁适口性好, 是家畜喜食的优良牧草[22], 而且由于其出苗成坪快, 建植性好, 质地柔软均一, 常和其他草种混播并被作为草坪建植的先锋草种[23]。不仅如此, “ 黑麦草-水稻(Oryza sativa)” 草田轮作系统在农业生产实践中产生了显著的经济效益, 黑麦草在国内外已经被广泛应用于生产实践, 发挥着巨大的生态和经济效应[24, 25, 26]。因此, 研究AM真菌和禾草内生真菌对黑麦草抗旱性的影响, 将有助于揭示干旱胁迫下, 两类微生物提高多年生黑麦草对水分利用效率的生理机制, 为黑麦草的广泛应用提供理论支撑。
目前, 国际上已有大量关于AM真菌以及禾草内生真菌与植物共生的研究报道, 二者单独作用对于植物生长、生理等方面的影响机制已逐渐清晰。但由于大多数报道都是基于单一微生物与宿主共生机制的研究, 忽略了两类微生物互作的潜在效应, 其所得结果具有一定的局限性[27]。现有的少数关于AM真菌和内生真菌互作对植物生长影响机制的研究中, 由于研究方法、微生物种类、宿主选择、试验条件等的差异, 得出的结论并不一致[28, 29]。在水分胁迫条件下, 二者互作对于彼此侵染植物的影响也有报道[30], 但具体机制尚需深入研究。本研究以多年生黑麦草为供试对象, 通过探究不同水分梯度下, 单独接种AM真菌或禾草内生真菌以及两类微生物互作对植物生长、养分吸收和抗旱性的影响, 以期为因地制宜合理利用两类微生物, 提高植物生长能力及抗旱性提供理论支撑。
1.1.1 供试植物 供试的宿主植物为多年生黑麦草, 种子AR1带禾草内生真菌(E+), Nil不带禾草内生真菌(E-), 由新西兰国家草地研究所(AgResearch)、新西兰皇家科学院院士John Caradus教授提供。播种前种子用10% H2O2表面消毒10 min, 无菌水冲洗干净后, 均匀摆放在铺有双层滤纸的直径100 mm培养皿内, 置于20~25 ℃恒温箱中催芽, 种子露白即可播种。
1.1.2 AM菌根真菌 AM真菌为摩西球囊霉(Glomus mosseae), 购买于北京市农林科学院植物营养与资源研究所, 编号为BGC NM04A。用红三叶(Trifolium pratente)扩繁所得的孢子及培养基质作为接种物。
1.1.3 供试土壤 供试土壤为砂土。将草炭土装在铁皮桶中, 121 ℃湿热灭菌1 h, 隔天再灭菌1 h, 自然风干; 将河沙过2 mm筛, 170 ℃干热灭菌6 h; 将兰州大学榆中校区黄土170 ℃干热灭菌6 h, 然后将草炭土、黄土和河沙以体积比1∶ 1∶ 2混匀, 密封备用。取20 g测定其营养含量及土壤pH。
混合土壤pH 7.6, 全磷含量19.60 mg· kg-1, 全氮含量18.38 mg· kg-1。
1.2.1 试验设计 盆栽试验于2016年3月26日-7月16日, 在兰州大学草地农业科技学院智能温室中进行。试验设置4种接菌处理:单接AM菌处理, 记作AME-; 单接禾草内生真菌处理, 记作NME+; 双接菌处理, 记作AME+; 不接菌处理, 记作NME-。每种接菌处理下按照田间持水量(field water content, FWC)的15%、30%、45%、60%共4个水分梯度进行干旱胁迫。每个处理设4个重复, 每个重复种植1盆(直径112 cm, 高15 cm), 共64盆多年生黑麦草。每盆从培养皿中随机移栽5株, 随机排列64个花盆, 隔天依次调换花盆位置, 使各盆所处条件一致, 出苗1周后, 选择长势一致的保留4株。
1.2.2 水分胁迫方法 黑麦草播种38 d后, 进行干旱胁迫。试验设4个土壤相对含水量梯度, 分别为田间持水量的15%、30%、45%、60%。按照4个水分梯度在第39天开始分梯度称量浇水, 每天最少称量一次, 天气较热时早晚各称量一次。具体称量标准:15%水分处理称量至1 093.78 g, 30%水分处理称量至1 136.60 g, 45%水分处理称量至1 179.43 g, 60%水分处理称量至1 222.26 g。其中, 盆重为50.95 g, 每盆土壤干重为1 000 g; 土壤最大持水量为285.51 g。此外, 温室温度保持在18~25 ℃、湿度保持在65%~80%、光照为自然光照, 室内开启内遮阴和空气加湿装置以最大程度减小蒸腾作用, 以保持花盆内的土壤含水量。
1.2.3 AM真菌接种方法 以穴播方式接种, 花盆装土时每盆先装土800 g, 接菌处理添加接种剂20 g, 不接种处理(NM)则添加等量灭活菌剂20 g, 以确保其微生物区系一致, 最后上层覆土180 g。
1.2.4 试验指标测定方法 AM菌根侵染率测定根据文献[31]的方法。
采用愈创木酚法[32]测定过氧化物酶(peroxidase, POD)活性, 采用硫代巴比妥酸(TBA)法[33]测定丙二醛(malondialdehyde, MDA)含量。
真菌对植株养分吸收量的贡献率[34]按照下式计算:
真菌对植株养分吸收量的贡献率=(接菌处理的吸收量-对照处理的吸收量)/接种处理的吸收量× 100%。
植物全氮和全磷含量测定:称取0.200 g干燥并研磨过的植物材料(干燥研磨过1 mm筛, 在70 ℃干燥3~4 h), 转移到1 000 mL消煮管中; 加入3.3 g(K2SO4∶ CuSO4=10∶ 1)催化剂和10 mL浓硫酸, 消煮至溶液澄清(溶液呈淡蓝绿色), 通常需要2 h; 冷却后定容至100 mL; 然后采用流动注射仪(FIAstar 5000 Analyzer, FOSS, Sweden)测定, 并计算植株的全氮、全磷含量和吸收量[35]。
土壤pH的测定:称取4份20 g充分混合均匀的土样, 在室温下风干, 置于50 mL的塑料离心管中, 加30 mL 0.01 mol· L-1CaCl2, 振荡1 h后用42号滤纸过滤溶液, 清液用酸度计测定其pH[36]。
1.2.5 试验数据统计 试验数据用Excel软件, 按照不同水分梯度和不同接菌处理进行整理汇总, 用JMP IN 4统计软件进行方差分析, 比较不同水分处理和接菌处理分别造成的差异, 通过多重比较法对差异显著的处理进行比对, 结果用GraphPad Prism 5.01作柱状图进行分析。
不接种AM真菌处理(NM)未检测到菌根结构, AM接菌处理菌根侵染率因不同水分条件和接菌处理而异。AM真菌、禾草内生真菌和水分三者的互作对菌根侵染率有极显著影响(P< 0.001)(表1)。在不同水分条件下AM真菌侵染率差异显著(P< 0.05), 其中45%水分条件下AM真菌侵染率最高, 平均达74.66%; 15%水分条件最低, 平均为35.05%。15%和60%水分条件下, AME-处理黑麦草根系的AM真菌侵染率较AME+处理分别高161.11%和53.28%(P< 0.05)。30%和45%水分条件下, AME-和AME+处理间的AM真菌侵染率无显著差异(P> 0.05)(图1)。
| 表1 水分、丛枝菌根及禾草内生真菌互作的多因素方差分析 Table 1 ANOVA results on the effects of water, mycorrhizal inoculation, grass endophyte treatment, and their interactions on the listed variables |
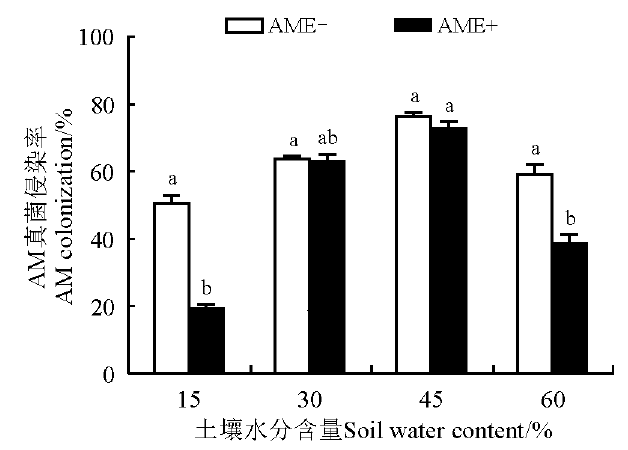 | 图1 不同土壤水分、AM真菌及禾草内生真菌条件下的AM真菌侵染率 图中不同小写字母表示同一水分条件下不同处理间存在显著差异(P< 0.05)。AME-, 单接AM 菌处理; AME+, 双接菌处理。Fig. 1 AM colonization on perennial ryegrass under different soil water treatments Different lowercase letters on the bars show significant differences between plants with the same water treatment at the 0.05 level. AME-, arbuscular mycorrhiza and non-endophyte; AME+, arbuscular mycorrhiza and endophyte. |
AM真菌与禾草内生真菌互作显著影响了各水分条件下植物生长(表1), 不同水分条件下黑麦草地上、地下生物量差异显著(P< 0.05), 地上生物量随土壤水分含量增加而增加(图2)。其中, 以15%水分条件下地上生物量最小, 60%水分条件下最大。在15%水分条件下, 两类共生真菌显著增加了地上生物量; 45%和60%水分条件下, 二者不同程度地增加了地上生物量。较两种共生菌单独作用, AM真菌和禾草内生真菌互作在60%水分条件下相较于AME-处理显著降低了黑麦草地上生物量, 其余水分条件下未显著影响地上生物量(P> 0.05)。
 | 图2 不同土壤水分、AM真菌及禾草内生真菌处理下的地上、地下生物量 图中不同大写字母表示不同水分间存在显著差异 (P< 0.05); 不同小写字母表示同一水分条件下不同处理间存在显著差异(P< 0.05); NME-, 不接菌处理; NME+, 单接禾草内生真菌; AME-, 单接AM菌处理; AME+, 双接菌处理处理; 下同。Fig. 2 Shoot and root biomass of E- and E+ perennial ryegrass inoculated with or without AM fungi under different soil water treatments Different capital letters indicate significant differences among plants treated with different water contents at the 0.05 level; different lowercase letters indicate significant differences among for the same water treatment different treatments at the 0.05 leve; NME-, non-mycorrhiza and endophyte; NME+, non-mycorrhiza and non-endophyte; AME-, arbuscular mycorrhiza and non-endophyte; AME+, arbuscular mycorrhiza and endophyte, similarly for the following figures. |
各接菌处理对黑麦草地下生物量的影响也随不同水分条件有所不同, 45%和60%水分条件下, AM真菌(AME-)与禾草内生真菌(NME+)较NME-分 别提高了27.5%~49.1%和10.1%~53.2%(P< 0.05); 而15%和30%水分条件下各接菌处理间差异不显著(P> 0.05)(图2)。
水分、AM真菌、禾草内生真菌三者的互作对黑麦草叶片相对含水量具有显著影响(P< 0.05), 水分对黑麦草叶片相对含水量具有极显著影响(P< 0.01)(表1)。随着土壤水分含量增加, 黑麦草叶片相对含水量总体呈递增趋势, 分别在15%和60%水分梯度下黑麦草叶片相对含水量最小和最大。15%水分条件下, AME-与NME+处理下黑麦草叶片相对含水量分别较NME-低12.58%和15.03%(P> 0.05)。在30%水分条件下, AME-与AME+处理下黑麦草叶片相对含水量较NME-分别下降了22.45%(P< 0.05)、17.56%。45%与60%水分条件下, 各接菌处理与NME-相比黑麦草叶片含水量无显著差异(P> 0.05)(图3)。
 | 图3 不同土壤水分、AM真菌及禾草内生真菌处理下的叶片和根系相对含水量Fig. 3 Leaf and root relative water contents of E- and E+ perennial ryegrass inoculated with or without AM fungi under different soil water treatments |
不同处理下, 根系相对含水量较为一致, 45%和60%水分条件下根系含水量低于其他处理(P< 0.05)(图3)。
水分处理、AM真菌和禾草内生真菌显著影响植物地上、地下N含量(P< 0.01)(表1)。接种AM真菌与禾草内生真菌均显著影响黑麦草地上部分全N 含量, 且接种处理的黑麦草地上部分全N含量显著高于不接种处理。在15%和45%水分条件下, E+(AME+和NME+)植株的全N含量较NME-有所提高但差异不显著(P> 0.05)(图4), AM真菌对植株养分吸收贡献率为负值, 且在15%水分梯度下差异显著(P< 0.05)(图5); 在30%水分条件下各接种处理与NME-的全N含量的差异均不显著(P> 0.05); 在45%水分条件下, 各接种处理均显著高于NME-(P< 0.05), 真菌对黑麦草N素吸收贡献率出现差异; 在60%水分条件下, 接种AM真菌处理的植株全N含量显著高于NME-, 禾草内生真菌处理则和NME-差异不显著(P> 0.05)。在15%和60%水分条件下, 两类微生物互作对黑麦草地上部分N素吸收贡献率高于其他接菌处理(图4、图5)。在45%和60%水分条件下, AME+处理显著提高了地下部分全N的含量, 且在不同水分梯度下各接菌处理间差异显著(P< 0.05)。
 | 图4 不同土壤水分、AM真菌及禾草内生真菌处理的全氮、全磷含量Fig. 4 Total nitrogen and phosphorus contents of E- and E+ perennial ryegrass inoculated with or without AM fungi under different soil water treatments |
 | 图5 不同土壤水分、AM真菌及禾草内生真菌处理下真菌对黑麦草氮、磷吸收的贡献率Fig. 5 Contribution of fungi to nitrogen and phosphorus uptake efficiency of E- and E+ perennial ryegrass inoculated with or without AM fungi under different soil water treatments |
与地上部分相同, 在15%和60%水分条件下, AM真菌与禾草内生真菌单独作用对地下部分全N含量并无显著影响(P> 0.05)。接种AM真菌在干旱条件下未能显著影响黑麦草地下部分全N含量, 而接种禾草内生真菌提高了黑麦草地下部分全N含量。除30%水分条件外, 其余水分条件下两类微生物互作均有效提高了对黑麦草根系N吸收的贡献率(图4、图5)。总体来看, 两类共生微生物在低水分条件下对黑麦草N吸收贡献作用不显著, 在45%和60%水分条件下则表现出显著促进作用。
在45%水分条件下, 接种AM真菌显著提高了黑麦草地下部分全P含量(P< 0.05); 在60%水分条件下, 与NME-相比, 各接菌处理均显著提高了黑麦草地下部分全P含量(图4、图5)。总体来看, 接菌处理对黑麦草地上部分P吸收起一定促进作用, 对于其地下部分P吸收贡献率则不明显。与其他接菌处理相比较, 在各水分梯度下, 两类微生物互作对于地上、地下P吸收贡献率无显著差异(P> 0.05)。
水分、AM真菌与禾草内生真菌对丙二醛含量、过氧化物酶活性(除水分× AM真菌)表现出显著的两两互作效应(P< 0.01)(表1)。AM真菌与禾草内生真菌在干旱条件下(15%)降低了丙二醛的含量, 与NME-相比, NME+、AME-、AME+处理下丙二醛含量降幅分别为43.68%、26.72%和27.11%(P> 0.05)。30%及45%水分条件下, 共生微生物表现出提高、降低或不影响丙二醛含量(图6)。在60%水分条件下, 各接菌处理与NME-之间差异不显著(P> 0.05)。各水分条件下, 以15%水分梯度下黑麦草丙二醛浓度最高。除15%水分梯度外, 其余水分条件下较单接菌处理, 两类微生物互作均提高了黑麦草丙二醛含量, 但差异不显著(P> 0.05)(图6)。
 | 图6 不同土壤水分、AM真菌及禾草内生真菌处理下的丙二醛浓度和过氧化物酶活性Fig. 6 MDA content and POD activity of E- and E+ perennial ryegrass inoculated with or without AM fungi under different soil water treatments |
过氧化物酶活性随土壤水分含量增加总体呈降低趋势。在15%和60%水分条件下, 两类共生微生物单独或共同作用均提高了植物POD酶含量, 与NME-相比, NME+、AME-、AME+处理在15%水分条件下POD活性增幅分别为34.83%、56.01%和16.70%, 在60%水分条件下, POD活性增幅依次为79.32%、137.97%和43.61%。30%和45%水分条件下, 各接种处理之间POD活性无差异, 但均显著高于NME-组(P< 0.05)。在15%和60%水分条件下, 两类微生物互作处理黑麦草POD活性显著低于AME-处理(P< 0.05)(图6)。
目前普遍认为, 禾草内生真菌和寄主的共生关系受到禾草内生真菌菌株类型[37]、宿主基因型[38, 39]、水分条件、养分条件、宿主生长环境[40]、二者的相互适应性[18]以及进化关系[14]等诸多因素的影响, 总体来看是一种基于互利共生和竞争拮抗的连续统一体[19]。唐明等[41]通过分析AM真菌对沙棘(Hippophae rhamnoides)抗旱性的影响后提出:一般情况下, 菌根内总菌丝量> 功能菌丝量> 活性菌丝量, 能够吸收利用养分的主要是具有磷酸酶活性的菌丝, 即AM真菌对植物营养吸收及抗旱性提高主要依靠具有磷酸酶活性的菌丝, 解释了AM真菌效应的差异。本研究中, AM真菌-禾草内生真菌-土壤水分存在互作关系, 且这种互作关系主要通过两类真菌在不同土壤水分条件刺激下, 表现出不同的活性及侵染优势[42], 从而对宿主的养分吸收及生理生化反应产生不同的影响。Staddon等[30]的研究表明, 干旱条件会通过刺激AM真菌和内生真菌孢子活性, 影响其初期菌丝延伸, 来调控两类微生物的侵染率, 但两类微生物相互之间没有发现相互调节侵染率的现象。本研究表明, 水分胁迫会调节两类微生物的侵染, 并且禾草内生真菌会抑制AM真菌的侵染[43, 44]。有研究指出, 禾草内生真菌菌丝体分布在禾草地上部分, 且幼嫩组织中菌丝密度较大, 随着植株组织成熟, 菌丝体密度减少, 在根系中未曾发现禾草内生真菌菌丝体[18, 20]。因此, 本研究推断禾草内生真菌是通过其产生的次生代谢产物抑制AM菌根的生长。禾草内生真菌与AM 真菌会竞争宿主所能提供的一定量的光合产物、水分、氧气等生存空间与资源。这与Heepper等[45]的结论类似。
本研究发现, 土壤水分含量充足时黑麦草植株生长迅速, 体内营养富集化程度较低, 干物质量以及营养元素含量较高。在不同水分条件下, 两类微生物均对黑麦草的生长有促进作用, 在15%水分条件下共生微生物对黑麦草地上部分和地下部分生长的促进程度不同, 在30%水分条件下共生微生物对黑麦草整株生长的促进作用不明显。两类共生微生物对黑麦草地上部分和地下部分养分吸收的贡献率均表现一致, 而且两类共生微生物对N吸收的贡献率较高, 对P吸收贡献率则出现负值。贺学礼等[46]研究发现, 不同水分条件下接种真菌对植株全N和全P的影响不同, 这和本研究的结论一致。总体来看, 在不同水分条件下, NME+处理对N的吸收保持较为稳定的促进作用; AME-处理则对P的吸收保持较为稳定的促进作用; 本研究中, 绝大多数条件下, 两类共生微生物互作对黑麦草养分吸收的贡献率为正, 且显著高于两类共生微生物单独作用时的贡献率, 因此认为, 两类微生物的互作效应也受到了土壤水分条件的刺激或调解。Omacini等[27]的研究也表明, AM真菌不管是否与禾草内生真菌互作, 都未提高植物的生物量及养分积累量或浓度。
MDA作为细胞膜脂的过氧化物, 是植物细胞膜脂过氧化程度的体现, 当植物细胞受到外界环境胁迫时, 植株体内丙二醛含量会显著升高, 从而刺激机体进行胁迫应答反应。本研究表明, 除15%水分条件外, 接种处理均不同程度地提高了黑麦草体内丙二醛含量。另外, 贺学礼等[47]的研究认为, POD活性在胁迫后期较高, 接种AM真菌可以显著提高POD活性, 而且AM真菌在植物生长前期的作用可以增强植物在生长中后期应对环境胁迫的能力。本研究中, 各接种处理下植物体内POD活性均显著提高, 而且各水分条件下植物POD活性处于一个相近的水平。推断植物缺水造成细胞膜受损, POD活性马上提高到一个临界水平来清除植物体内的过氧化物自由基, 而且接菌处理可以极大程度地提高这个临界水平。这和已有的禾草内生真菌可以显著提高植物POD活性的报道一致[48]。本研究表明, 接菌处理并没有显著影响植株叶片相对含水量, 只是相对改善了黑麦草水分分配效率。Elmi和West的研究[39]表明:禾草内生真菌对于高羊茅(Festuca arundinacea)植物细胞维持生存能力的影响远大于其促进植物生长能力的影响。而且有学者认为, AM菌丝网吸收的水分并不多, 只够向根系提供一部分水分而无法满足植物地上部分对水分的需求[49, 50], 这些都和本研究所得结论一致。
综上, AM真菌与禾草内生真菌仅在45%和60%最大田间持水量条件下, 显著促进了黑麦草生长以及养分吸收, 并以两种菌互作的效果更为显著, 在更为干旱的条件下, 两类微生物并未有效促进植物生长, 这表明, AM真菌与禾草内生真菌互作, 仅在一定土壤水分条件下对多年生黑麦草有促生作用。
(责任编辑 武艳培)
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|