

{kind=link}
{kind=link}
{kind=link}
{kind=link}
土地利用方式对豫西黄土丘陵区土壤团聚体微生物生物量及群落组成的影响
[刘晶 , 张跃伟, 张巧明, 徐少君]
, 张跃伟, 张巧明, 徐少君]
, 张跃伟, 张巧明, 徐少君]
|
|


采用熏蒸浸提和磷脂脂肪酸(PLFA)法研究了豫西黄土丘陵区不同土地利用方式下土壤团聚体中微生物生物量及其群落组成。结果表明,0.25~2 mm粒级团聚体微生物生物量碳(MBC)和微生物生物量氮(MBN)含量最高,0.053~0.25 mm粒级团聚体MBC和MBN含量最低;阔杂林、刺槐林和灌草地>5、2~5和0.25~2 mm 3个粒级团聚体的MBC和MBN含量较耕地显著提高( P<0.05),果园与耕地之间无显著差异( P>0.05)。不同利用方式下,细菌PLFA含量及其占团聚体总PLFA的比例分别以0.25~2和0.053~0.25 mm粒级团聚体最高;真菌PLFA含量及其占比为>5 mm粒级团聚体最高,且随团聚体粒径的减小而降低。MBC和MBN含量与PLFA总量、表征细菌和真菌PLFA含量均显著或极显著正相关( P<0.05或 P<0.01)。与耕地和果园土壤相比较,阔杂林地、刺槐林地和灌草地中>5、2~5和0.25~2 mm粒级团聚体中表征真菌PLFA的含量显著增加。综上,阔杂林、刺槐林和灌草地较耕地和果园显著提高了>0.25 mm粒级团聚中微生物生物量及真菌含量。
Microbial biomass and community composition in aggregates of different land use types in the loess hill region of west Henan were analyzed by fumigation extraction method and phospholipids fatty acids (PLFA) biomarkers. Microbial biomass carbon (MBC) and microbial biomass nitrogen (MBN) contents were the highest in aggregate of 0.25~2.00 mm and lowest in 0.053~0.250 mm. Compared with the farmland, the MBC and MBN contents in >5 mm, 2~5 mm and 0.25~2.00 mm aggregates of broad-leaved mixed forestland, Robinia pseudoacacia plantation forestland and shrub-grass land were significantly increased( P<0.05), which in orchard and farmland were no significant difference( P>0.05). The PLFA contents of bacteria were the highest in the 0.25~2.00 mm aggregates, however, the highest proportion of bacteria to total PLFA were found in 0.053~0.250 mm aggregates in all different land use types. The PLFA contents of fungi and its proportion to total PLFA in the >5 mm aggregates were the highest and induced with the decrease of the aggregates size. The contents of MBC and MBN present a notable positive correlation with the PLFA contents of the bacteria and fungi( P<0.05 or P<0.01). Compared with the farmland and orchard soil, the contents of fungi in>5 mm, 2~5 mm and 0.25~2.00 mm aggregates of broad-leaved mixed forestland, R. pseudoacacia plantation forestland and shrub-grass land increased significantly. To sum up, soil microbial biomass and fungi PLFA contents in >0.25 mm aggregates of broad-leaved mixed forestland, R. pseudoacacia plantation forestland and shrub-grass land were significantly increased compared with farmland and orchard.
土壤微生物是土壤中物质转化和养分循环的驱动力, 对土壤有机质的分解和理化性质的改善起着重要作用[1]。土壤团聚体是微生物的重要栖息场所, 也是土壤的重要结构单元, 其形成和稳定性都与微生物关系密切。团聚体的数量和质量直接决定着土壤的质量和肥力[2, 3]。不同粒级土壤团聚体的物理和化学特性各异, 导致其微生物类群和活性有很大不同, 从而导致不同粒级土壤团聚体物质循环和能量转化的速度存在显著差异[4, 5]。土壤微生物生物量作为土壤中物质代谢强度的指标, 能够较早地指示生态系统功能的变化, 反映人类活动对土地利用方式和生态功能的影响[6]。国内对不同耕作、施肥措施、植被恢复和土地利用变化等对全土微生物生物量和群落结构的影响方面的研究较多。玉米(Zea mays)连作、玉米非连作和撂荒3种不同种植方式下, 土壤微生物量和群落组成有明显的差异[7]。化肥与有机肥、化肥与豆科绿肥配合使用均不同程度提高了茶园土壤微生物生物量碳、氮含量及土壤酶活性[8]。岩溶槽谷区不同用地类型土壤中有机质、全钾、有效钾和有效磷的含量与土壤微生物之间的相互关系显著[9]。施加生物碳和凋落物能有效提高与氮素循环相关的微生物数量, 改变土壤中细菌与真菌的比例[10]。但现有对土壤微生物生物量和群落结构的研究大多集中在原土水平上, 缺乏对田间土壤过程的客观反映。本研究采用干筛法筛分豫西黄土丘陵区不同利用方式地土壤团聚体, 对不同粒级团聚体中微生物生物量和群落组成进行初步分析, 其结果有助于揭示微生物与土壤团聚体稳定性之间的相互作用, 也能进一步解释不同土地利用类型下植被演替过程及其人为干扰对土壤质量的作用机制。
研究样地位于洛宁县(34° 05'-34° 38' N、111° 08'-111° 49' E)境内, 2006-2015年年均降水量550 mm, 年均气温14.2 ℃, 土壤类型为棕壤。选择5类不同利用类型样地, 阔杂林(B)为天然生栓皮栎(Quercus variabilis)和麻栎(Quercus acutissima)混交林; 刺槐林(R)为人工栽种8年刺槐(Robinia pseucdoacaia); 灌草(S)地主要植被为酸枣(Ziziphus jujuba ‘ spinosa’ )、伴生蒿类(Artemisia spp.)和芒草(Miscanthus sinensis)等草本植物; 果园(O)栽种苹果(Malus pumila); 耕地(F)种植小麦(Triticum aestivum)。阔杂林地和刺槐林地林下植被多, 湿度大, 果园和耕地由于人为管理, 地表基本无杂草。采样方法及土样预处理参照刘晶等[11]方法进行。
湿筛法和干筛法是分离土壤团聚体的两种常用方法。湿筛法由于采用水浸泡等过程, 易使团聚体中微生物生物量的分布发生变化。而干筛法可以相对减少对微生物群落的破坏, 在旱作条件下, 干筛法分离出的团聚体更接近田间实际[12], 故本研究中采用干筛法分离土壤团聚体。取5、2、0.25和0.053 mm的筛子, 按孔径大小从上到下组成一组套筛, 称取200 g风干土样平铺于最上层筛子上, 将盛有土样的套筛固定在水平震荡机上后以200 r· min-1速度筛分3 min, 收集各层筛面上的团聚体并称重记录, 分别为> 5, 2~5, 0.25~2, 0.053~0.25 mm粒级的团聚体, 重复两次。将收集的各粒径团聚体过0.25 mm筛、分别称取各粒级团聚体样品30 g, 调节湿度至田间持水量的50%后置于50 mL小烧杯中, 在4 ℃冰箱中平衡24 h, 置于25 ℃恒温培养箱中培养7 d(不同利用方式地每一粒级团聚体培养设置6个重复), 其中3个样品用于测定微生物生物量碳、氮, 3个样品在-20 ℃下保存, 用于PLFAs的测定。
微生物生物量采用氯仿熏蒸-浸提法[13]。微生物生物量碳含量采用重铬酸钾氧化外加热法[14], 全氮含量采用半微量凯氏法[14], 转化系数取0.45[15]。土壤微生物磷脂脂肪酸提取参照修正的Bligh-Dyer方法[16, 17]。
脂肪酸种类通过脂肪酸Sherlock微生物鉴定系统(MIDIInc., Newark, DE, USA)进行鉴定, 采用Frostegard方法命名。磷脂脂肪酸和土壤微生物的对应关系如表1所列[18, 19, 20]。
| 表1 磷脂脂肪酸与土壤微生物生物量的对应关系 Table 1 Phospholipid fatty acids associated with different soil microbes |
所有数据经Excel整理后, 用SPSS 18.0软件进行方差分析, 不同利用方式之间多重比较及其显著性水平(P< 0.05)通过最小显著差数法LSD进行检验。
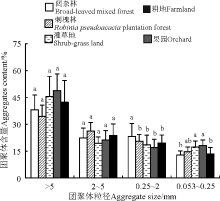
5种土地利用方式均以> 5 mm土壤团聚体含量最高, 2~5 mm团聚体含量次之, 0.053~0.25 mm 团聚体含量最低, > 2 mm团聚体占团聚体总量的60%以上, 各粒级团聚体含量总体呈现出随粒径减小而降低的趋势(图1)。与耕地相比较, 阔杂林地、刺槐林地、灌草地和果园土壤> 5和2~5 mm团聚体含量均无显著变化(P> 0.05); 阔杂林地0.25~2 mm团聚体含量显著高于其他地(P< 0.05), 灌草地和果园土壤0.053~0.25 mm粒级土壤团聚体含量也显著高于阔杂林和耕地(P< 0.05)。
 | 图1 不同利用方式地土壤团聚体含量不同小写字母表示同一团聚粒径不同利用方式间之间差异显著(P< 0.05), 下同。Fig. 1 The soil aggregates content of different land use soilsDifferent lowercase letters indicate significant difference among different land use types of same aggregate size at the 0.05 level; similary for the following figures. |
微生物生物量碳(MBC)含量为233.25~964.21 g· kg-1, 微生物生物量氮(MBN)含量为73.32~24.64 g· kg-1(图2)。各粒级团聚体MBC和MBN含量均以阔杂林地最高, 耕地最低。各利用方式地中, 阔杂林地全部4个粒级团聚体MBC含量均为最高, 较耕地分别显著提高了146.17%、152.83%、135.10%和111.07%, MBN含量分别显著提高了135.88%、112.28%、89.32%和72.61%(P< 0.05)。刺槐林地和灌草地各粒级团聚体MBC、MBN含量也均显著高于耕地(P< 0.05)。果园仅0.053~0.25 mm粒级团聚体MBC、MBN含量显著高于耕地(P< 0.05)。同一利用方式地0.25~2 mm粒级土壤团聚体MBC含量最高, 2~5 mm粒级次之, 阔杂林、刺槐林、果园和耕地0.25~2和2~5 mm粒级团聚体MBC、MBN含量均显著高于> 5和0.053~0.25 mm粒级(P< 0.05)。
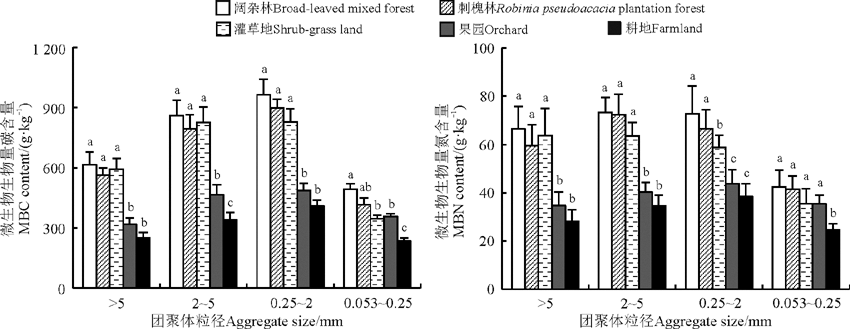 | 图2 不同粒级团聚体中土壤微生物生物量碳、氮含量Fig. 2 Contents of MBC and MBN in different size aggregates |
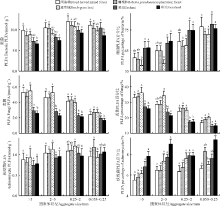
2.3.1 磷脂脂肪酸(PLFA)含量及比例 不同利用方式地各粒级土壤团聚体表征细菌、真菌和放线菌的PLFA含量分别为6.62~11.92、1.69~5.28和0.78~1.21 nmol· g-1(图3)。阔杂林地、刺槐林地和灌草地各粒级团聚体中表征细菌和真菌的PLFA含量较耕地均显著提高(P< 0.05), 其中阔杂林地0.25~2 mm粒级团聚体表征细菌的PLFA含量最高, 为11.92 nmol· g-1, 灌草地> 5 mm粒级团聚体中表征真菌的PLFA含量最高, 为5.48 nmol· g-1, 分别较耕地含量显著提高了29.14%和112.40% (P< 0.05), 提高幅度最大; 果园土壤中0.053~0.25 mm粒级团聚体表征细菌的磷脂脂肪酸含量较耕地含量显著提高了21.42%(P< 0.05)。灌草地0.25~2 mm粒级团聚体表征放线菌的PLFA含量显著高于耕地 (P< 0.05), 其他均与耕地之间无显著差异(P> 0.05)。同一利用方式地不同粒级团聚体中, 表征细菌的PLFA含量以0.25~2 mm 团聚体最高, 0.053~0.25 mm团聚体次之, > 5 mm团聚体最低; 真菌PLFA含量> 5 mm团聚体最高, 2~5 mm团聚体次之, 0.053~0.25 mm团聚体最低。不同利用方式地各粒级团聚体中表征细菌的PLFA含量占PLFA总量的57.77%~76.63%, 真菌为36.10%~14.62%, 放线菌为5.50%~9.43%。同一利用方式地, 0.053~0.25和> 5 mm粒级团聚体中表征细菌和真菌的PLFA分别在PLFA总量中占比最高。
 | 图3 不同粒径团聚体中磷脂脂肪酸含量及比例Fig. 3 Contents and rations of PLFA in different size aggregates |
2.3.2 磷脂脂肪酸含量与微生物生物量碳、微生物生物量氮关系 MBC含量与PLFA总量、表征细菌和革兰氏阴性菌的PLFA含量表现为极显著正相关关系(P< 0.01), 与表征真菌的PLFA含量呈显著正相关关系(P< 0.05), 与表征革兰氏阳性菌的PLFA含量呈显著负相关系(P< 0.05), 与表征放线菌的PLFA含量相关性不显著(P> 0.05)(表2)。MBN含量与PLFA总量、表征细菌、真菌和革兰氏阴性菌的PLFA含量都表现为极显著的正相关着重(P< 0.01), 与表征放线菌和革兰氏阳性菌的PLFA含量相关性不显著(P> 0.05)。
| 表2 微生物生物量碳、微生物生物量氮与各磷脂脂肪酸标记的相关系数 Table 2 The correlation coefficients between the MBC, MBN and the PLFA |
2.3.3 不同粒级团聚体中磷脂脂肪酸主成分分析 对各利用方式地不同粒级团聚体中PLFA进行了主成分分析(图4)。不同粒径从大到小第一主成分的贡献率分别为42.23%、47.68%、51.81%和38.44%, 第二主成分的贡献率分别为26.22%、31.36%、28.96%和21.86%, PC1的贡献率普遍高于PC2。进一步对不同利用方式各粒级团聚体在PC1和PC2上的得分系数进行方差分析(表5)。从PC1分析, 除0.053~0.25 mm粒径团聚体外, 阔杂林地、刺槐林地、灌草地3种利用方式地与果园和耕地2种利用方式在其余3个粒级均存在显著差异(P< 0.05); 从PC2分析, 在> 5和0.25~2 mm粒组中, 阔杂林地、刺槐林地、灌草地3种利用方式与果园和耕地存在显著差异(P< 0.05)。由此可见, 与耕地和果园土壤相比较, 阔杂林地、刺槐林地和灌草地 3种利用方式显著改变了> 5、2~5和0.25~2 mm粒级团聚体中微生物的群落组成。
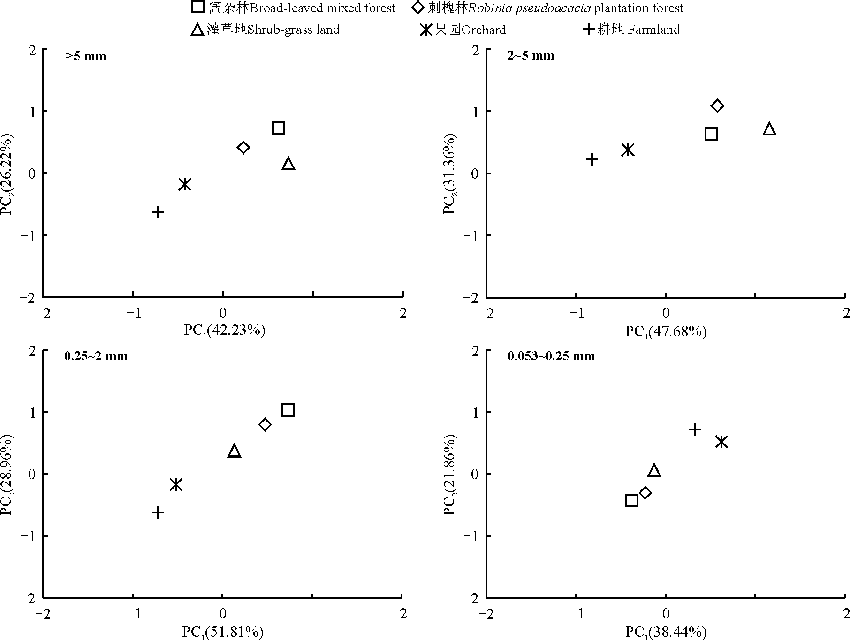 | 图4 不同粒径团聚体中微生物PLFA主成分分析Fig. 4 PCA of microorganism PLFA in different size aggregate |
| 表3 不同利用方式地各粒径团聚体微生物得分系数分析 Table 3 The PCA scores of microorganisms in different size aggregates in different land use type |
土壤微生物生物量的变化能够灵敏地反映土地利用和管理上的差异, 常作为不同利用方式或耕作措施引起土壤生物学性质变化的指标之一[21]。土壤微生物生物量碳和微生物生物量氮作为土壤养分中比较活跃的部分, 对土壤扰动的反应比较敏感, 其分布特征可以在一定程度上表征土壤的质量与肥力[4, 22]。退耕等生态恢复措施可以显著提高全土中微生物生物量含量[21, 23, 24], 本研究中, MBC和MBN含量均以0.25~2 mm粒级团聚体最高, 2~5 mm粒级次之, 0.053~0.25 mm粒级最低, 基本按土壤团聚体粒径从小到大呈现中间高两边低的变化趋势。在几种利用方式地中, 0.25~2和2~5 mm两个粒级团聚体的MBC、MBN含量分别占到团聚体MBC、MBN总量的60%和55%以上。主成分分析也表明, 0.053~0.25 mm粒级团聚体中PC1和PC2均比较小, 表明它的PLFA含量较低, 也就是土壤所含微生物较少。可见, 黄土丘陵区中等大小的团聚体中土壤微生物生物量最高, 粒径过大或者过小不利于土壤微生物的生存。该结果与国外部分研究结果[25, 26, 27]及国内对亚热带地区紫色水稻土[28]、喀斯特峰丛洼地区[29]不同粒级团聚体中MBC分布情况有所不同。一方面土壤微生物量在不同粒径的团聚体中的分布可能与团聚体的孔径差异有关, 土壤微生物生物量与土壤孔隙度呈显著的指数正相关关系[30]; 另一方面这可能与不同粒径团聚体中有机质的分解程度有关, 由于粒径较大的土壤团聚体透气性好, 碳循环周转速率较快, 微生物难以保证足够的养分来维持其生长与繁殖。细菌是土壤中多样性最丰富的微生物类群, 也是土壤营养物质循环的主要执行者之一, 在土壤养分循环中起着至关重要的作用, 且其数量和类群变化可以及时反映土壤质量和生态环境的变化情况[31, 32, 33, 34]。本研究结果显示, 在不同土地利用方式下, 各粒级团聚体中细菌PLFA占团聚体微生物总量的80%以上, 细菌PLFA含量均远高于真菌群落, 说明细菌群落在该区土壤微生物中占据主要地位。同一利用方式地不同粒径土壤团聚体间细菌生物量变化较大, 但不同利用模式地各粒径团聚体中细菌生物量分布模式相似, 几种利用方式地都以0.25~2 mm粒级团聚体细菌生物量为最高。Chiu等[35]研究表明, 在亚高山草原和森林土壤颗粒中, > 0.25和0.002~0.053 mm粒级团聚体中细菌的生物量比0.053~0.25 mm粒级团聚体中高。这说明团聚体内部微生物群落结构有较大的变异性, 细菌在粒径较小的团聚体中占优势, 其原因可能是因为细菌可以在更小孔隙微团聚体生存更长时间。
真菌在团聚体的形成和稳定性等方面起着重要作用[36]。真菌菌丝体既可以起到绊缠等物理作用, 还可以通过分泌多糖类物质, 使土壤团聚体粘结在一起形成稳定的大团聚体[7, 37]。不同土地利用方式会导致土壤结构的变化, 进而影响不同粒级团聚体中真菌群落的组成, 数量及分布[2]。本研究显示, 阔杂林地、刺槐林地和灌草地团聚体真菌PLFA总量较果园和耕地土壤显著提高(P< 0.05), 阔杂林地、刺槐林地和灌草地三者之间差异不显著(P> 0.05), 果园和耕地两者之间差异也未达显著水平(P> 0.05)。各利用方式地土壤团聚体中真菌PLFA含量随粒径的递减而递减。由于不同利用方式地的植被差异很大, 这就使得植物根系和返还到土壤中的凋落物存在很大不同。植物根系在生长发育过程中, 不断释放化学物质, 这些根系分泌物或根际沉积物作为重要能源和养分源, 影响根际微生物的生长繁殖[22]。凋落物物理化学性状不一样, 如碳氮比和木质素含量等, 在分解过程中释放的有机无机物有很大的差异, 不同的分解物质可能对土壤微生物的生长具有刺激或抑制作用, 同时地表植物多样性的提高能够增加根际分泌物的种类和菌根的形成, 有助于提高土壤中真菌的数量及多样性[38, 39]。阔杂林地、刺槐林地和灌草地土壤团聚体中真菌生物量较果园和耕地土壤高, 可能是因为这几种利用方式下, 地表植被生物量和多样性较高, 返还到土壤中的凋落物数量增加, 凋落物的分解物质为真菌的生长提供了较为充足的养分, 促进了土壤真菌的繁殖生长; 或者是这几类利用地植物根系分泌物中某些成分更利于真菌的生长繁殖; 同时土壤机械扰动少, 能保持稳定的土壤孔隙, 更利于真菌菌丝的延展。团聚体中微生物群落的差异则可能是因为返还到土壤中的凋落物和根系分泌物在不同粒级团聚体中分配存在差异, 从而引起不同粒级团聚体中有机碳的结构组成和生物可利用性不同而造成的。对研究区不同利用方式地植被凋落物生物量、化学组成、在团聚体中的分配及其分解过程还待进一步研究。阔杂林地、刺槐林地和灌草地> 0.25 mm粒级团聚体与果园和耕地土壤在PC1和PC2均产生显著差异, 说明较大粒组的土壤团聚体中微生物群落组成对土地利用变化的响应更灵敏。
微生物是土壤营养物质循环的主要执行者之一, 不同的微生物种群在物质循环中执行着不同的任务。磷脂脂肪酸分析虽能够定量描述环境样品中的微生物群体, 适合用作微生物群落的总体分析, 但该方法尚不能从菌种和菌株水平精确分析环境中微生物的种类, 因而无法进一步解释微生物多样性的变化情况。因此, 采用分子生物学技术探析团聚体中微生物群落菌种的组成与结构变化应该是下一步研究的重点。
1)不同利用方式地土壤团聚体中, > 2 mm团聚体占团聚体总量的60%以上, 各粒级团聚体含量随粒径减小而降低, 几种利用方式地土壤> 5和2~5 mm团聚体含量均无显著差异(P> 0.05)。
2)不同利用方式地各粒级土壤团聚体MBC和MBN均以0.25~2 mm粒级团聚体最高, 2~5 mm粒级次之, 0.053~0.25 mm粒级最低。阔杂林、刺槐林和灌草地> 5、2~5和0.25~2 mm 3个粒级MBC和MBN含量较耕地显著提高(P< 0.05), 果园与耕地之间无显著差异(P> 0.05)。
3)不同利用方式地各粒级土壤团聚体中, 细菌占微生物总量的57.77%~76.63%, 真菌为36.10%~14.62%, 放线菌生物量最少; 同一利用方式地, 细菌生物量在0.25~2 mm粒级团聚体中最高, 而真菌生物量则在> 5 mm粒级团聚体中最高。
4)MBC和MBN 与PLFA总量、表征细菌和真菌的PLFA含量均显著正相关(P< 0.05或P< 0.01)。阔杂林地、刺槐林地和灌草地> 5、2~5和0.25~2 mm粒级团聚体中真菌生物量较果园和耕地显著增加(P< 0.05)。
(责任编辑 苟燕妮)
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|