

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
高寒草甸湿润与干旱化生境中植物多样性的海拔分布格局
[张大才 , 李双智, 张锐]
, 李双智, 张锐]
, 李双智, 张锐]
|
|


受全球变化和外界干扰的影响,青藏高原高寒草甸生境趋向干旱化。生境变化对物种多样性的海拔分布格局有何影响?本研究以藏东南东达山作为研究地点,沿海拔梯度设置9个样地,在每个样地设置10个1 m×1 m样方,5个位于湿润生境,5个位于干旱生境,调查样方内的物种组成。将样方中频度≤7的物种确定为偶见种,反之为常见种,分析所有种、偶见种和常见种α、β和γ多样性沿海拔梯度的分布格局。以Cody指数(βC)、倍性分配法(βW)和物种组成相似性随海拔距离的衰减速率(βslope)3种方法测度β多样性。样地中共记录种子植物160种,其中湿润生境132种,干旱生境107种。湿润生境中,所有种和偶见种的α和γ多样性随海拔的增加而下降,而干旱生境中呈现为增加趋势;常见种α和γ多样性在两种生境中均随海拔的增加而略增加。3种β多样性指数的分布格局不一致,湿润生境中βC和βW多样性高于干旱生境,偶见种βC和βW多样性高于常见种。高寒草甸湿润与干旱生境物种多样性的海拔分布格局不同,偶见种多样性高,直接影响物种多样性的海拔分布格局。
Because of the influence of global change and external disturbances, habitats of alpine meadows are trending toward aridification in the Qinghai-Tibetan Plateau. How would variation in habitat affect the elevational pattern of species diversity? Nine sample plots were set along an elevational gradient in the Dongda Mountains in southeast Tibet. Ten 1 m×1 m sample quadrats were set in each sample plot, with five quadrats located in wet habitat and the others located in dry habitat. Species composition in each quadrat was investigated. Species with a frequency lower than seven were defined as occasional species; otherwise, the species were common. For total occasional and common species, variation of α, β, and γ diversity were respectively analyzed along the elevational gradient. β diversity was measured by the Cody index (βc), multiplicative partitioning (βw), and the slope of regression between similarity of species composition and distance (βslope). There were 160 seed plants recorded in these sample plots, among which, 132 species were distributed in wet habitats and 107 species in dry habitats. For total and occasional species, α and γ diversity both decreased with elevational gain in wet habitats but increased in dry habitats. For common species, α and γ diversity both presented an increasing trend with elevational gain in wet and dry habitats. The elevational patterns of β indexes were not consistent,βc and βw in wet habitats were higher than that in dry habitats, and βc and βw of occasional species was higher than that of common species. The elevational patterns of species diversity between wet and dry habitats were different, and diversity of occasional species had a high value, and will directly affect elevational patterns of species diversity.
物种多样性空间分布格局是生物多样性保护和生物地理单元划分的基础, 一直受到高度关注[1, 2]。生境受到外界干扰后, 物种组成和多样性也随之改变[3], 生境变化是否影响物种多样性的分布格局?高寒草甸是青藏高原分布面积最大的植被类型之一, 受全球变化和外界干扰严重[4, 5], 生境趋向干旱化, 群落退化演替, 物种多样性下降[6]。藏东南高寒草甸在生境干旱化过程中, 植物群落仍然以嵩草属(Kobresia)植物为优势种, 根据土壤水分条件可分为湿润与干旱化两种生境类型[7]。生境类型与物种多样性密切相关, 生境的变化都将引起植物群落类型和物种多样性的变化, 如不同坡向高寒草甸物种多样性不同[8], 林中空地、沟谷、湿地等不同生境物种多样性存在差异[9]。但是, 更多研究都主要比较不同生境类型间物种多样性的差异, 而很少分析不同生境物种多样性分布格局的差异, 本研究将比较藏东南高寒草甸湿润与干旱化生境植物多样性沿海拔梯度分布格局的差异。
在外界干扰过程中, 高寒草甸群落结构发生了很大的改变[10, 11, 12], 一些物种的个体形态趋向小型化[13], 植物体构件数量明显下降[14], 在群落中的分布频度低而成为群落中的偶见种(occasional species)。同时, 植物群落受到外界干扰后, 改变了群落物种组成和种间关系, 为其他物种侵入群落提供了机会[15], 这些侵入性物种也可能是分布频度低的偶见种。藏东南高寒草甸受放牧干扰严重, 群落中偶见种对物种多样性组成及海拔分布格局有何影响?本研究将植物群落中的物种区分为偶见种和常见种, 比较其多样性沿海拔梯度的分布格局。
α 多样性和γ 多样性统称编目多样性, 测度物种组成多样性, 但二者测度的尺度不同, α 多样性测度局域尺度上的物种多样性, 而γ 多样性测度区域尺度上的物种多样性[16]。β 多样性测度环境梯度上物种组成的变化, 其值对α 和γ 多样性起关键作用。β 多样性的测度方法有多种, 如倍性分配法、加性分配法、物种组成相似性等, 然而众多方法的测度结果不完全一致[17, 18]。β 多样性概念的提出是为了反映物种组成沿环境梯度的变化[19], 物种的分布频度必将影响β 多样性, 分布频度低的偶见种对β 多样性的贡献更高[20, 21, 22]。γ 多样性在更大空间尺度上测度群落物种多样性, 与α 和β 多样性的分布格局是否具有一致性?因此, 本研究主要研究目的有:1)分析高寒草甸植物群落种子植物多样性沿海拔梯度的分布格局; 2)比较高寒草甸湿润与干旱化生境中物种多样性沿海拔梯度分布格局的差异; 3)分析偶见种对物种多样性组成的贡献。
研究地点位于藏东南左贡县东达山, 该山山体高大, 垭口海拔5 008 m。研究区域地貌以高山为主, 地形呈“ 〕 ” 形, 沟谷地形平坦, 宽度约200 m, 溪流纵贯其间, 生境湿润, 地表稍有积水或湿润; 两侧沿山体斜升, 生境趋向干旱化, 地表干燥。整个区域均处于放牧干扰状态, 无围栏区域, 放牧强度几乎一致。研究区域属高原半干旱气候, 气温年较差小, 年平均气温为0 ℃左右, 最暖月平均气温为10 ℃以下; 降水集中于夏季, 冬季干燥寒冷[23]。
东达山高寒草甸以嵩草属植物为优势种, 分布于林线与高山流石滩之间, 分布的海拔范围近1 000 m。从林线边缘的高寒草甸开始设置样地, 海拔每升高约100 m设置一个样地, 直至高山流石滩边缘, 共9个样地, 除1、2号样地外, 其余样地坡度相差不大(表1)。在每个样地设置10个1 m× 1 m样方, 其中第1-5号为湿润生境样方, 设置于沟谷溪边地表稍有积水或土壤表面湿润的生境中; 第6-10号样方为干旱化生境(干旱生境)样方, 均设置于北坡, 地表干燥; 第10号样方生境极其干旱, 为嵩草属植物的分布边缘, 样方外为高山灌丛。调查样方内的物种组成、盖度和多度等调查因子。
| 表1 样地信息 Table 1 Information of sample plots |
1.3.1 偶见种的确定 偶见种指在群落中出现频度低的物种, 从研究的实际出发, 有多种确定方法, 如将出现在所有样地的物种确定为常见种, 反之则为偶见种[24]; 或者根据物种在群落中出现的频度确定, 将频度低于一定标准的物种确定为偶见种[21, 22, 25]; 或者根据研究者的经验判断, 将个体数量少的物种确定为偶见种[26]。本研究根据物种在样方中出现的频度确定群落中的偶见种, 将频度低于群落中物种平均频度的物种作为偶见种。湿润和干旱生境中物种在样方中出现的平均频度分别为6.8和7.1, 因此本研究中将频度≤ 7的物种作为偶见种, 将频度> 7的物种作为常见种, 分析所有种、偶见种和常见种多样性沿海拔梯度的分布格局。
1.3.2 物种多样性的测度 α 多样性:以样方内物种数量测度α 多样性, 即物种丰富度。γ 多样性:为区域性物种多样性, 即一个大的区域内的物种数量, 本研究在海拔梯度上共设置9个样地, 以每个海拔带湿润和干旱生境所有样方记录的物种总数分别测度该海拔样地湿润和干旱生境的γ 多样性。β 多样性:1) Cody指数, β C=[g(H)+l(H)]/2, g(H)为沿海拔梯度增加的物种数, l(H)为沿海拔梯度减少的物种数[27]。2)倍性分配法, β W=γ /
1.3.3 数据统计分析 采用SPSS 16.0进行回归分析, 分析物种多样性与海拔梯度之间的回归关系, 并绘制回归关系图。使用ANOVA方差分析比较湿润与干旱两种生境中物种多样性的差异, 用离散系数(coefficient of variance, CV)衡量同一样地不同样方间物种丰富度分布的均匀程度, CV值越大表示分布越不均匀, CV值为0则表明各样方物种丰富度相同, 分布最均匀, CV=标准差/平均值。
东达山90个样方中共记录种子植物160种, 其中湿润生境132种, 干旱生境107种, 两种生境物种组成Simpson相似性系数为0.736。湿润生境中有偶见种71种, 占湿润生境物种总数的68.9%; 干旱生境中有偶见种75种, 占干旱生境物种总数的70.1%。
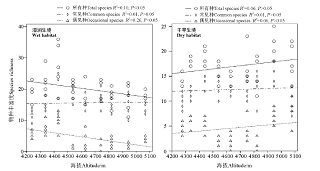
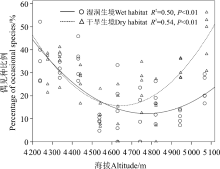
湿润生境中各样方所有种、常见种丰富度显著高于干旱生境(P< 0.05), 而偶见种丰富度在两种生境中的差异不显著(P> 0.05)(表2)。湿润生境中, 所有种和偶见种丰富度随海拔的升高而呈显著线性下降(P< 0.05), 而常见种丰富度随海拔的增加略呈增加趋势(P> 0.05)(图1)。干旱生境中, 所有种、偶见种和常见种丰富度随海拔的增加均呈现出增加趋势(P> 0.05)(图1)。常见种丰富度在两种生境中随海拔梯度增加的增幅均较小, 且远远低于所有种和偶见种; 湿润生境中所有种和偶见种丰富度沿海拔梯度的变化速率也高于干旱生境(表2)。偶见种丰富度占所有种丰富度的百分比沿海拔梯度的变化呈现为单峰曲线格局, 且最低值出现在中海拔段, 即低、高海拔段偶见种所占的比例最高(图2)。同一样地内不同样方间物种丰富度差异明显, 离散系数(CV)变化范围为0.05~1.02。湿润生境CV值高于干旱生境; 两种生境中, CV值均表现为偶见种> 所有种> 常见种。
| 表2 湿润与干旱生境物种多样性比较 Table 2 The comparison of plant species diversity between wet and dry habitats |
 | 图1 物种丰富度沿海拔梯度的分布格局Fig. 1 Patterns of plant species richness along an elevational gradient |
 | 图2 湿润与干旱生境中偶见种丰富度占所有种丰富度比例沿海拔梯度的变化Fig. 2 Percentage of occasional species richness to total species richness varied along an elevational gradient |
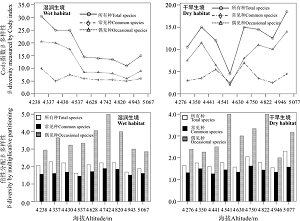
湿润生境中, β c多样性随海拔的升高而下降(图3), 而干旱生境中, β c多样性在中海拔区域最低(图3); 两种生境中, β w多样性总体上在中海拔区域最高(图3)。除湿润生境偶见种外, 物种组成相似性与海拔高差之间均呈显著线性相关关系, 偶见种物种组成相似性最低, 而常见种物种组成相似性最高(图4)。两种生境中偶见种β C、β W多样性均显著高于常见种(P< 0.05), 且在9个样地中偶见种β W多样性始终高于常见种; 但是, 偶见种β slope多样性低于常见种; 湿润生境β C和β W多样性高于干旱生境, 而β slope多样性低于干旱生境(表2)。
 | 图3 β 多样性沿海拔梯度的变化Fig. 3 Variation of beta diversity along an elevational gradient |
 | 图4 Simpson相似指数与海拔高差之间的线性回归关系Fig. 4 Linear regression between Simpson similar index and relative altitude |
湿润生境中所有种和偶见种γ 多样性随海拔增加而下降, 而常见种γ 多样性随海拔的增加而增加; 干旱生境中所有种、常见种、偶见种γ 多样性均随海拔的增加而增加; 偶见种γ 多样性高于常见种γ 多样性, 且常见种γ 多样性在两种生境中随海拔增加的增幅很小, 基本保持不变(表2, 图5)。
 | 图5 γ 多样性沿海拔梯度的变化Fig. 5 Variation of Gamma diversity along an elevational gradient |
东达山高寒草甸编目多样性沿海拔梯度的分布格局表现出3个特征。第一, 常见种、偶见种多样性格局差异明显。偶见种与所有种编目多样性沿海拔梯度的分布格局几乎一致(表2, 图1、5), 说明偶见种对群落物种编目多样性的海拔分布格局起着重要作用。高寒草甸植物群落的优势种大多以克隆繁殖为主[30], 这些优势种也是群落的常见种, 它们的海拔分布范围大, 物种编目多样性沿海拔梯度的变化小。同时, 偶见种占所有种比例在高寒草甸分布的低、高海拔段最高, 即在高寒草甸与森林、与高山流石滩的生态交错区最高(图2)。不同植被类型交错区植物多样性常常都很高[31],
而且物种相互作用机制表明部分物种维持最低生物量以保持群落物种的多样性[32], 这与偶见种分布频度低、个体数量少的分布特征一致, 表明偶见种是生态交错区物种多样性的重要组成部分。第二, 编目多样性与海拔梯度的相关性不高。同一样地不同样方间物种丰富度差异明显(图1), 丰富度离散程度大, 离散系数最大值超过1, 且偶见种丰富度的离散程度最大。相邻样方间物种组成及多样性差异大, 导致多样性与海拔梯度之间的相关性不高, 与东灵山[33]、小五台山[34]亚高山草甸植物多样性沿海拔梯度的分布格局一致。因此, 如果取样强度不够也可能导致物种丰富度沿海拔梯度分布格局的假相, 如很多研究表明高寒草甸植物群落物种丰富度沿海拔梯度的变化呈现“ 单峰” 曲线格局[35, 36]。无论从地质历史、植被分布、气候因子、物种分布几何边界的中间膨胀效应等各个方面, 似乎都无法解释这一分布格局。第三, 湿润与干旱生境物种多样性沿海拔梯度的分布格局差异明显, 两种生境中编目多样性基本呈现为相反的变化趋势(表2, 图1、5)。在海拔梯度上编目多样性主要有两种分布格局, 一是随海拔的增加而下降的偏峰格局, 另一种是随海拔的升高先增加而后下降的单峰曲线格局[37]。但是, 不管哪种分布格局, 在高海拔段物种丰富度都一致地表现为随海拔的升高而下降的偏峰格局, 尤其在高大的山体[38, 39, 40, 41, 42]。湿润生境中所有种和偶见种编目多样性沿海拔梯度的分布格局符合这一普遍现象, 而干旱生境物种编目多样性与这一普遍分布格局相反。物种多样性沿海拔梯度的分布格局受多种因子的影响, 如气候、土壤、植被、生境类型等[43]。本研究中, 湿润与干旱生境位置相邻, 气候、环境与植被因子沿海拔梯度的变化相似, 但物种多样性的海拔分布格局不同, 说明两种生境植物群落对外界干扰的抵抗力不同, 干旱生境植物群落及物种多样性受外界干扰严重, 物种多样性的分布格局已完全改变。大量针对草地开展的研究都表明物种多样性与群落稳定性之间呈现正相关关系, 高物种多样性有利于维持群落稳定性[44, 45, 46]。高寒草甸干旱生境物种多样性低, 群落稳定性低, 生境干旱化过程是植物群落退化演替的过程。
3种β 多样性测度方法的结果不完全一致(图3、4), 其中, β C和β W结果表明, 偶见种β 多样性高于常见种(图3), 而β slope结果表明, 偶见种β 多样性低于常见种(图4)。偶见种在群落中的分布频率低, 在环境梯度上的替换速率快, 对群落β 多样性贡献更高[20, 21, 22], 与本研究中β C和β W的测度结果一致。β C和β W均为常见的β 多样性测度方法, 有广泛的使用, 其测度结果可靠性高, 而β slope指数使用相对较少, 既不属于β 多样性的倍性测度方法, 也不属于β 多样性的加性测度方法, 其测度结果有待于进一步检验。同时, β C和β W多样性指数在海拔上的变化也不完全相同, 偶见种β W多样性始终高于常见种, 而对于β C而言, 偶见种β C多样性大多数情况下高于常见种, 而在一个海拔样地上偶见种β C多样性低于常见种(图3)。因此, β W多样性指数能够更好地区分群落物种的组成, 对群落物种组成的变化更加敏感。
高寒草甸植物群落中偶见种多样性高, 海拔分布格局与所有种一致, 直接影响群落物种多样性和海拔分布格局。高寒草甸干旱生境植物群落物种多样性更低, 与多样性沿海拔梯度分布的普遍格局相反, 是退化演替植物群落物种多样性的分布特征。偶见种在植物群落中的分布频度低, 在环境梯度上的替代率高, 对β 多样性贡献高, 且对其变化敏感, 其结果可以反映不同β 多样性指数的测度效果。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|